ИЗМЕНЕНИЕ КОРТИКО-СПИНАЛЬНЫХ МЕХАНИЗМОВ РЕГУЛЯЦИИ ИЗОМЕТРИЧЕСКОГО МЫШЕЧНОГО НАПРЯЖЕНИЯ СПОРТСМЕНОВ В УСЛОВИЯХ ЭКСТРЕМАЛЬНЫХ НАГРУЗОК
Работа выполнена при поддержке Министерства образования и науки РФ (НК-116П/6, П444)
ФОМИН Роман Николаевич
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Заведующий лабораторией нервно-мышечной физиологии НИИ спорта РГУКФСиТ, к.б.н., доцент
E-mail: RomanFominVL@gmail.com, тел: +7-916-639-79-80
СЕРГЕЕВ Виталий Николаевич
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Лаборатория нервно-мышечной физиологии НИИ спорта РГУКФСиТ, мнс.
E- mail: niinst@mail.ru, тел. +7-985-960-99-90
ЖЕРНОСЕК Анна Михайловна
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Руководитель НОЦИ РГУКФСиТ, к.п.н.
E- mail: anna528@mail.ru, тел. +7-916-691-86-41
ЧЕРЯТНИКОВ Александр Юрьевич
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Кафедра теории и методики бокса и кикбоксинга, аспирант.
E-mail: sanya-807@mail.ru, тел: +7-926-288-61-48
СЕЛЯЕВ Михаил Васильевич
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Лаборатория нервно-мышечной физиологии НИИ спорта, м.н.с.
E-mail: vaselyaev@rambler.ru, тел: +7-926-516-60-35
КОСМЫНИН Владимир Сергеевич
Российский государственный университет физической культуры, спорта, молодёжи и туризма (ГЦОЛИФК), Москва
Лаборант кафедры АФК
E- mail: kosmyninvladimir@inbox.ru, тел. +7-962-939-31-18
Ключевые слова: кортико-спинальные механизмы, изометрическое мышечное сокращение, экстремальная двигательная деятельность, магнитная стимуляция.
Аннотация. При помощи метода транскраниальной магнитной стимуляции нервно-мышечного аппарата человека исследованы адаптивные изменения кортико-спинальных механизмов управления изометрическим мышечным сокращением спортсменов в условиях экстремальных нагрузок.
CHANGE IN THE CORTICO-SPINAL MECHANISMS OF REGULATION OF ISOMETRIC MUSCLE TENSION ATHLETES UNDER EXTREME LOADS
This work was supported by the Ministry of Education and Science (NC-116P / 6, P444)
FOMIN Roman
Russian State University of Physical Culture, Sports, Youth and Tourism (GTSOLIFK), Moscow
Head of Laboratory of Neuromuscular Physiology, Institute of Sport ГЦОЛИФК, PhD, Associate Professor
E-mail: RomanFominVL@gmail.com, tel. 7-916-639-79-80SERGEEV Vitaliy
Russian State University of Physical Culture, Sports, Youth and Tourism (GTSOLIFK), Moscow
Laboratory of Neuromuscular Physiology, Institute of Sport RGUKFSiT, junior.
E-mail: niinst@mail.ru, tel. 7-985-960-99-90ZHARNASEK Anna
Russian State University of Physical Culture, Sports, Youth and Tourism (GTSOLIFK), Moscow
Head Nozi RGUKFS&T, PhD
E-mail: anna528@mail.ru, tel. 7-916-691-86-41SELYAEV Mikhail
Russian State University of Physical Culture, Sport and Tourism (GTSOLIFK)
Laboratory of Neuromuscular Physiology, Institute of Sport, Junior
E-mail: vaselyaev@rambler.ru, tel: +7-926-516-60-35CHERYATNIKOV Alexander
Russian State University of Physical Culture, Sport and Tourism (GTSOLIFK)
Department of theory and techniques of boxing and kickboxing, a graduate student.
E-mail: sanya-807@mail.ru, tel: +7-926-288-61-48KOSMYNIN Vladimir
Russian State University of Physical Culture, Sports, Youth and Tourism (GTSOLIFK), Moscow
Laboratory Assistant Department of AFC
E-mail: kosmyninvladimir@inbox.ru, tel. 7-962-939-31-18Keywords: cortico-spinal mechanisms, isometric muscle contraction, extreme motor activity, magnetic stimulation.
Summary. Using the method of transcranial magnetic stimulation of the neuro-muscular can man investigate the adaptive changes of cortico-spinal control mechanisms isometric muscular contraction athletes under extreme loads.
Актуальность. Одной из важнейших проблем физиологии спорта является исследование механизмов и закономерностей процесса адаптации организма к различным условиям среды [5,7,9]. Адаптация к экстремальной двигательной деятельности в процессе многолетней спортивной подготовки, обеспечивающей рост мастерства спортсмена, представляет собой очень сложный многоуровневый процесс, затрагивающий нейрональные механизмы управления адаптивными реакциями [7,9]. С ростом спортивной квалификации механизмы управления движениями человека становятся более совершенными, возрастают функциональные возможности организма, улучшается координация движений и т.п. [3,1].
В связи с этим, представляло интерес изучение состояния кортикальных и спинальных механизмов управления мышечным сокращением при выполнении изометрического мышечного усилия различной величины и длительности у спортсменов, адаптированных к экстремальной двигательной деятельности и различающихся уровнем спортивного мастерства. Мы предположили: чем выше уровень спортивной квалификации, тем более спортсмен адаптирован к экстремальной двигательной деятельности, а кортико-спинальные механизмы управления мышечным сокращением в процессе выполнения изометрических напряжений будут у него изменяться в зависимости от уровня адаптации.
Цель. Изучить изменения кортикальных и спинальных механизмов регуляции изометрического мышечного сокращения у спортсменов, адаптированных к экстремальной двигательной деятельности.
Испытуемые. В исследовании приняли участие 30 практически здоровых спортсменов-мужчин от 18 до 25 лет, специализирующихся в спортивных единоборствах (дзюдо, греко-римская борьба и бокс). Обследуемые были условно разделены на контрольную группу (15 низкоквалифицированных спортсменов, не адаптированных к экстремальной двигательной деятельности, - КГ) и экспериментальную группу (15 высококвалифицированных спортсменов, адаптированных к экстремальной двигательной деятельности, - ЭГ).
Организация и методы исследования. Исследование проводилось в лаборатории нервно-мышечной физиологии НИИ спорта РГУФКСиТ в 2010 г. Стандартный протокол исследования был таким: в начале обследования испытуемые посредством подошвенного сгибания стопы выполняли 3 максимальных произвольных сокращения (МПС) с интервалом отдыха 1 мин; затем последовательно выполняли произвольные изометрические мышечные напряжения, равные 25, 50, 75 и 100% от МПС. В процессе данных нагрузок применялись транскраниальная магнитная стимуляция (ТМС) моторной коры головного мозга, магнитная стимуляция (МС) сегментов спинного мозга и периферического нерва. При этом в покое, а также на 1, 15-й и 30-й секундах изометрических мышечных напряжений измеряли контрлатеральные вызванные моторные ответы (ВМО) скелетных мышц нижних конечностей, время центрального моторного проведения (ВЦМП), корковый и периферический периоды "молчания" электромиограммы (ЭМГ). Дополнительно регистрировались электрические ответы скелетных мышц нижних конечностей (М-ответ, Н-рефлекс) и период молчания ЭМГ при электрической стимуляции афферентных и эфферентных волокон периферического нерва.
Регистрация параметров произвольного и вызванного мышечного сокращения. С целью получения фоновых, текущих и итоговых данных о показателях произвольных и вызванных сокращений скелетных мышц спортсменов в изометрических условиях, а также моделирования нагрузки при выполнении мышечных сокращений использовался мультисуставной лечебно-диагностический комплекс "Biodex Multi-Joint System Pro-4" (Biodex Medical System, USA). Изометрическое мышечное сокращение, составляющее 25, 50, 75 и 100% от МПС, выполнялось в положении сидя посредством подошвенного сгибания стопы в условиях фиксации голени и коленного сустава. Тестирование предусматривало 3 предварительных сокращения стопой в упоре устройства и 3 последующих МПС. Периоды отдыха составляли 1 мин.
Магнитная стимуляция нервно-мышечного аппарата на фоне произвольных изометрических сокращений применялась последовательно: сначала стимулировали головной мозг, затем - спинной, в заключение - периферический нерв (рис. 1). ТМС моторной коры головного мозга осуществлялась при помощи магнитного стимулятора "Magstim Rapid" (Magstim Company, UK) через двойную катушку диаметром 110 мм, расположенную в точке пересечения вертекса и линии, соединяющей наружные слуховые проходы. При этом подбирали такое положение катушки, при котором ВМО скелетных мышц имели постоянную амплитуду и форму [6,10]. МС сегментов спинного мозга осуществлялась через плоскую катушку с внешним диаметром 70 мм, расположенную на уровне шейного и поясничного отделов позвоночника [6,10]. МС периферического нерва наносилась через сдвоенную катушку (бабочка) с внешним диаметром 25 мм, расположенную в области подколенной ямки. Интервалы отдыха между различными видами МС составляли не менее 5 мин.
Поверхностная электромиография осуществлялись после МС по общепринятой методике [2,4] с помощью электромиографического комплекса "Нейро-МВП-8" (ООО "Нейрософт", Россия). Электроды располагались на 4 скелетных мышцах голени: m. soleus (SOL), m. gastrocnemius mediale (GM), m. gastrocnemius laterale (GL) и m. tibialis anterior (TA).
Электростимуляция периферического нерва. Н- и М-ответы SOL вызывались по общепринятой методике путем стимуляции n. tibialis при помощи электростимулятора "Нейро-МВП-8" (ООО "Нейрософт", Россия) через биполярный электрод, расположенный в подколенной ямке. Анализировались величина порога, латентный период, амплитуда, длительность и форма [2].
Период молчания ЭМГ [13], характеризующий тормозные механизмы в нервно-мышечном аппарате, регистрировали при ТМС моторной коры (корковый период молчания), МС поясничного сегмента спинного мозга на уровне L5-S1 и n. tibialis, а также электрической стимуляции n. tibialis (периферические периоды молчания). Период молчания ЭМГ при всех видах стимуляции измерялся от конца ВМО, зарегистрированного на фоне изометрического мышечного, до конца абсолютного биоэлектрического молчания [4,13]. Сила магнитной индукции, необходимая для регистрации периода молчания ЭМГ, выражалась в виде коэффициента *1,25, на который умножалось значение порога контр-ВМО.
Результаты исследования и их обсуждение. Исследование кортико-спинальных механизмов управления изометрическим мышечным сокращением у лиц, адаптированных к экстремальной двигательной деятельности, выявило существенные различия в изменениях исследуемых показателей в процессе выполнения статических напряжений разной величины и длительности по сравнению с лицами, не адаптированными к экстремальной двигательной деятельности.
Результаты исследования кортикальных механизмов при ТМС. Максимальная амплитуда ВМО скелетных мышц SOL, GM, GL и TA по мере повышения величины и длительности статических усилий у адаптированных спортсменов возрастала более выраженно по сравнению с неадаптированными спортсменами (рис. 2). Это указывает на более выраженное усиление нисходящих потоков со стороны коры головного мозга в процессе увеличения изометрического сокращения у лиц, адаптированных к экстремальной двигательной деятельности.
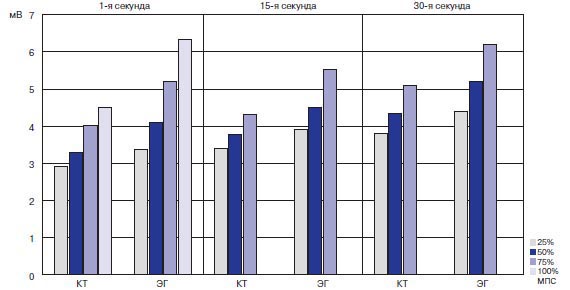
Рис. 2. Среднегрупповые значения максимальной амплитуды ВМО SOL в КГ и ЭГ при ТМС головного мозга на фоне статического усилия разной величины и длительности
Время центрального моторного проведения нервных импульсов от коры головного мозга до спинномозговых корешков у лиц, адаптированных к экстремальной двигательной деятельности, сокращалось в процессе изометрического мышечного сокращения в меньшей степени, чем у неадаптированными лиц. Так, в состоянии относительного мышечного покоя ВЦМП SOL в ЭГ составляла 20 мс. При ТМС моторной коры головного мозга амплитуда ВЦМП SOL на 1-й секунде изометрического сокращения при 25% от МПС составила 19,7 мс, при 50% -19,6 мс, при 75 % - 19 мс и при 100% - 18,6 мс. Указанные абсолютные значения были достоверно ниже величин ВЦМП SOL, зарегистрированных в состоянии покоя на 2, 2, 5 и 7% соответственно (p<0,05). Кроме того, результаты в ЭГ при 25, 50, 75 и 100% от МПС на 1-й секунде удержания превышали аналогичные показатели в КГ на 5, 8, 9 и 13% (p<0,05). Аналогичная динамика ВЦМП при ТМС моторной коры зарегистрирована в ЭГ на 1, 15 и 30-й секундах удержания. Возможно, это объясняется меньшим влиянием интракортикального торможения со стороны управляющих структур головного мозга на спинальные структуры лиц, адаптированных к экстремальной двигательной работе, что подтверждается собственными результатами при регистрации периода молчания ЭМГ.
Амплитудный коэффициент ВМО/М-ответ (АКм) SOL, GM, GL и TA у лиц, адаптированных к экстремальной двигательной деятельности, по сравнению с неадаптированными лицами, повышается менее выражено в процессе изометрического мышечного сокращения. Такая динамика связана с менее выраженным снижением М-ответа при повышающемся ВМО, что свидетельствует о более выраженном влиянии периферических мотонейронов на поддержание необходимого усилия по сравнению с неадаптированными лицами. Так, в состоянии относительного мышечного покоя АКм SOL в ЭГ составлял 15%. В условиях ТМС моторной коры головного мозга АКм SOL на 1-й секунде изометрического сокращения при 25% от МПС составил 30%, при 50% достиг значения 43%, при 75% увеличился до 76%, а при МПС повысился до 102% (p<0,05). Результаты АКм в ЭГ при 25, 50 и 75% от МПС на 1-й секунде удержания были ниже прироста аналогичных показателей в КГ на 25, 113 и 243% соответственно (p<0,05) и, напротив, на 80% выше АКм в КГ при 100% от МПС (p<0,05). Аналогичная динамика АКм при ТМС моторной коры зарегистрирована на 1, 15 и 30-й секундах удержания мышц-синергистов - GM и GL, а также у антагониста - TA.
На наш взгляд, это можно объяснить избирательным влиянием специфической адаптации к экстремальной двигательной деятельности на корковые и спинальные мотонейроны спортсменов. В процессе многолетних тренировок кортико-спинальные механизмы управления мышечным сокращением высококвалифицированных спортсменов становятся более приспособленными к повышающимся нагрузкам и модулируются более эффективно: при увеличении статических усилий в большей мере используется прямой моторный контроль мышечного сокращения.
Амплитудный коэффициент ВМО/H-ответ SOL, GM, GL и TA у лиц адаптированных к экстремальной двигательной деятельности, по сравнению с неадаптированными лицами имеет однонаправленную повышающуюся динамику в процессе выполнения изометрического мышечного сокращения. Это говорит о менее значимой роли рефлекторных механизмов поддержания изометрического сокращения у данных лиц.
Результаты исследования спинальных и периферических механизмов при МС. При МС поясничного сегмента спинного мозга на уровне L5-S1 в процессе изометрических сокращений разной величины и длительности у лиц, адаптированных к экстремальной двигательной деятельности установлены следующие факты:
• Максимальная амплитуда ВМО в мышцах SOL, GM, GL и TA у лиц адаптированных к экстремальной двигательной деятельности, по сравнению с неадаптированными лицами возрастает менее выраженно по мере повышения величины и длительности статических усилий. Это указывает на менее сильное возрастание эфферентных влияний не только со стороны моторной коры головного мозга, но и со стороны спинного мозга по мере повышения статических усилий у лиц, адаптированных к экстремальной двигательной деятельности.
• Время моторного проведения нервного импульса от спинного мозга до мышц SOL, GM, GL и TA у лиц, адаптированных к экстремальной двигательной деятельности, уменьшается менее выраженно в ходе повышения величины и длительности изометрического мышечного усилия. Аналогичная динамика наблюдалась при ТМС моторной коры головного мозга.
• Амплитудный коэффициент ВМО/М-ответ SOL, GM, GL и TA по мере повышения величины и длительности изометрического мышечного сокращения у лиц, адаптированных к экстремальной двигательной деятельности, повышается менее выражено, по сравнению с неадаптированными лицами. Это происходит за счёт меньшего снижения М-ответа при повышающемся ВМО скелетных мышц на фоне МС поясничного сегмента спинного мозга на уровне L5-S1: ослабление кортикального влияния и усиление спинального менее выражено в группе адаптированных к экстремальной деятельности. В процессе изометрических сокращений разной величины и длительности у лиц, адаптированных к экстремальной двигательной деятельности, обнаружена менее выраженная понижающаяся антагонистическая динамика результатов максимальной амплитуды ВМО мышц SOL, GM, GL и TA по сравнению с данными, полученными при ТМС моторной коры и МС поясничного сегмента спинного мозга на уровне L5-S1, у лиц, не адаптированных к экстремальной деятельности. Исследуемый показатель менее выражено понижался по мере повышения величины и длительности статических усилий.
Изменение тормозных кортико-спинальных механизмов у лиц, адаптированных к экстремальной двигательной деятельности, менее выражено в зависимости от величины и длительности изометрических сокращений. Корковый и сегментарный периоды "молчания" ЭМГ мышц SOL, GM, GL и TA в процессе выполнения изометрического мышечного сокращения разной величины и длительности у лиц, адаптированных к экстремальной двигательной деятельности, увеличиваются менее выражено по мере возрастания статического напряжения по сравнению с неадаптированными лицами. Это свидетельствует о менее выраженном усилении активности кортикальных и спинальных тормозных механизмов ЦНС, направленных на снижение биоэлектрической активности работающих мышц. Напротив, периферический период молчания ЭМГ мышц SOL, GM, GL и TA у лиц, адаптированных к экстремальной двигательной деятельности, понижается менее значительно по мере возрастания статического напряжения.
Таким образом, с одновременным усилением активности кортикальных и спинальных тормозных механизмов ЦНС, направленных на понижение электроактивности скелетных мышц на фоне нарастающего утомления, происходит менее выраженное растормаживание периферических структур нервно-мышечного аппарата у лиц, адаптированных к экстремальной двигательной деятельности, по сравнению с неадаптированными лицами. Это подтверждается другими исследованиями, в которых установлено, что в развитии периода "молчания" принимают участие спинальные и корковые механизмы подавления активности мотонейронов: в первой трети периода "молчания" (около 50 мс) задействована комбинация корковых и спинальных механизмов (рефрактерность спинальных мотонейронов и возвратное торможение клеток Реншоу), но все остальное время этого периода обусловлено корковым подавлением [8].
Литература
1. Бернштейн Н.А. Очерки по физиологии движений и физиологической активности / Н.А. Бернштейн. - М.: Медицина. - 1966. - 349 с.
2. Городничев Р.М. Спортивная электронейромиография / Р.М. Городничев. - Великие Луки: ВЛГАФК, 2005. - 229 с.
3. Козловская И.Б. Афферентный контроль произвольных движений / И.Б. Козловская. - М., 1976. - 296 с.
4. Команцев В.Н. Методические основы клинической электронейромиографии / В.Н. Команцев, В.А. Заболотных. - СПб., 2001. - 350 с.
5. Меерсон Ф.З. Адаптация к стрессовым ситуациям и физическим нагрузкам / Ф.З. Меерсон, М.Г. Пшенникова. - М.: Медицина, 1988. - 254 с.
6. Никитин С.С. Магнитная стимуляция в диагностике и лечении болезней нервной системы. Руководство для врачей / С.С. Никитин, А.Л. Куренков. - М.: САШКО, 2003. - 378 с.
7. Павлов С.Е. Адаптация / С.Е. Павлов. - М.: Паруса, 2000. - 282 с.
8. Персон Р.С. Спинальные механизмы управления мышечным сокращением / Р.С. Персон. - М.: Наука, 1985. - 184 с.
9. Платонов В.Н. Адаптация в спорте / В.Н. Платонов. - Киев: Здоровья, 1988. - 216 с.
10. Barker A.T., Jalinous R. And Freeston I.L. Non-invasive magnetic stimulation of human motor cortex. // Lancet. - 1985. - V.1. - P.1106-1107.
11. Chen R., Lozano A.M., Ashby P. Mechanism of the silent period following transcranial magnetic stimulation. Evidence from epidural recordings. Exp. Brain Res. 1999; 128(4): 539-543.
12. Garvey M.A., Ziemann U., Becker D.A., Barker C.A., Bartko J.J. New graphical method to measure silent periods evoked by transcranial magnetic stimulation. Clin. Neurophysiol. 2001; 112: 1451-1460.
13. Inghilleri M., Berardelli A., Crucci G., Manfredi M. Silent period evoked by transcranial stimulation of the human cortex and cervicomedullary junction. J. Physiol. 1993; 161: 112-125.