Abstract TOWARDS PROBLEM OF TERMINOLOGY IN REHABILITOLOGY V.A. Kachesov, scientific worker Yu.B. Mikhaylova, post graduate St.-Petersburg state P.F. Lesgaft academy of physical culture Key words: function, life, vialibity, vital activity, biological death, regeneration. The aim of this study was to substantiate the definitions of "function", "life", "biological death", "regeneration of tissues" in the aspect of rehabilitology's problems. The authors had offered these definitions and concluded that the clear comprehension of the dialectic unity of structure and function as well as all the above mentioned notions will allow rehabilitologists to avoid the useless discussions and the pessimistic forecasts about the reversibility and irreversibility of pathological states and the distroyed functions' rehabilitation.
|

К ВОПРОСУ О ТЕРМИНОЛОГИИ В РЕАБИЛИТОЛОГИИ Научный сотрудник В.А.
Качесов Аспирантка Ю.Б. Михайлова Ключевые слова: функция, жизнь, жизнеспособность, жизнедеятельность, биологическая смерть, регенерация. Любая научная дисциплина базируется на четком понятийном аппарате. В реабилитологии одно из основных понятий - функция, так как восстановление функции является основной задачей реабилитологов. И хотя о единстве структуры и функции говорил еще Р. Декарт, до сих пор нет четкого определения, связывающего два этих понятия. Образно о структуре и функции высказался известный терапевт В.Х. Василенко: "Функция без структуры немыслима, а структура без функции бессмысленна" [12]. Обобщая дискуссионный материал, Д.Т. Саркисов дает такое определение функции: "Биологическая функция - это деятельность, т.е. изменение во времени и пространстве состояния или свойств тех или иных структур организма и его самого как целого" [12]. Здесь же он замечает, что взаимоотношения структуры и функции до сих пор являются предметом острейших дискуссий [12]. Авторы данной статьи дают свое определение функции, которое объединяет понятия структуры и функции. Для этого необходимо рассмотреть процессы сокращения и расслабления гладкомышечного волокна, как наиболее изученные на данном этапе развития науки. От способности мышечных клеток функционировать зависят в конечном итоге гомеостаз и жизнедеятельность всего организма. Механизм и последовательность реакций мышечного сокращения доказаны также наблюдениями с помощью электронного микроскопа. Рассматривая вопрос о том, живет организм или нет, мы руководствуемся комплексом признаков, а в конечном итоге - оценкой мышечных сокращений поперечно-полосатой и гладкой мускулатуры (передвижение организма, дыхание, сердцебиение и т.д.). Гладкая мускулатура широко представлена в человеческом организме циркулярными волокнами во всех трубчатых органах (сосуды, кишечник, бронхи, трахея, протоки желез и каналов, желчный и мочевой пузыри, зрачок), актин, миозин или их комплекс содержатся во всех клетках для осуществления митоза, амебовидного движения, фагоцитоза, секреции [17, 19]. Чтобы показать механизм действия гладкомышечных волокон, рассмотрим процесс сокращения (сжатия) и расслабления одного из них. Фаза сокращения (синтеза актомиозинового комплекса). Если мышечная клетка не сжата и не перерастянута, то такое состояние называется состоянием покоя. В этот момент клеточная мембрана поляризова на, а клетка готова совершить работу [10]. Механизм синаптической передачи в холинэргических синапсах заключается в том, что при выделении ацетилхолина (АХ) в нейромышечном синапсе возбуждается холинорецептор, происходит резкое изменение ионной проницаемости и возникает потенциал действия (ПД). В результате происходящей деполяризации мембраны изменяется электрическое поле, которое открывает в мембране натриевые каналы. В клинической практике по изменению электромагнитного поля определяют специфическую функцию органа ( ЭКГ, ЭЭГ и т.д.). После возникновения ПД через короткий промежуток времени может произойти сокращение мышечного волокна за счет движения актина и миозина внутриклеточных миофибрилл относительно друг друга. В момент возбуждения миофибриллы ее мембрана становится проницаемой для ионов Са++, который, войдя в клетку, активизирует миозин. В процессе сокращения важную роль играет цГМФ ( циклический нуклеотид - вторичное сигнальное вещество). Рецепторы, расположенные на внешней поверхности клетки, связываются с лигандами, что сопровождается активацией мембранной олигоферментной системы - гуанилатциклазы, необходимой для модуляции 3,5- цГМФ. Реакция идет в присутствии ионов Са++ [11]. (Модуляция цГМФ и цАМФ необходима как энергетически выгодный процесс для создания этих универсальных катализаторов и вызываемого затем при их участии каскада реакций, происходящих при сокращении и расслаблении с минимальными затратами энергии [9, 11, 20].) Расход энергии макроэргов (ГТФ и креатин-фосфата) будет соответствовать вводимому количеству Са++. При этом действует фермент Са++Мg++АТФаза. Свойства миозиновой АТФазы таковы, что она попеременно действует то как Са++, то как К+Мg++Са++АТФаза, то есть участвует в сокращении и расслаблении [11, 14, 17, 18]. Таким образом, проявление специфической функции клетки, в данном случае - сокращения, обязательно сопровождается следующими процессами: выход К+ из клетки, вход Nа+ в клетку, вход Са++ в клетку, гидролиз трифосфатов, модуляция цГМФ и выделение энергии. Резко возрастает потребление кислорода. Поляризация мембраны исчезает - происходит деполяризация, затем возникает ПД, подчиняющийся закону "все или ничего" и, наконец, осуществляется собственно сокращение - синтез актомиозинового комплекса [14, 16]. Остановка сокращения (синтеза актомиози нового комплекса). Остановка гидролиза ГТФ и других макроэргов в случае гипотетической остановки сокращения гладкомышечного волокна за счет модуляции 3,5-цАМФ и других механизмов заставляет каскад реакций мгновенно выводить продукты метаболизма (СО2 и Н2О и др.), в результате чего метаболический ацидоз не нарастает [11, 14, 16-18]. Фаза расслабления ( распада актомиозинового комплекса). После процесса сокращения гладкомышечного волокна и наступления контрактуры необходима сила (каскад биохимических реакций), ведущая к расслаблению мышцы - распаду актомиози нового комплекса. Этот процесс возникает при возбуждении адренорецептора медиатором симпатином - смесью норадреналина и адреналина [17]. Адренорецептор, связанный через лигандный комплекс с аденилат-циклазой, модулирует 3,5-цАМФ, и начинается каскад реакций, приводящих к распаду актомиозинового комплекса. В этот момент снова действует универсальный фермент К++ МГ++ АТФаза. Са++, NА+, СL- выводятся из клетки, выводятся также окончательные продукты метаболизма (СО2, Н2О и др.) [ 8]. В клетку вводятся К+, и энергетические субстраты: глюкоза и т.д. Происходит накопление макроэргов (АТФ, ГТФ, креатин-фосфата), т.е. потенциальной энергии [8, 9, 10, 17]. Состояние покоя. Для мышц, находящихся в состоянии покоя и не расходующих энергии, характерна очень низкая скорость дыхания, т.е. потребления кислорода. В этих условиях концентрация АТФ и ГТФ высокая, а АДФ и ГДФ - низкая [8, 9]. Активные центры актина и миозина заблокированы ионами К+, и они представляют собой две раздельные конформации белков [16, 18]. Если рассматривать секрецию как специфическую функцию, то она обусловлена теми же процессами, что и мышечное сокращение [14, 17, 19]. Для выброса секрета нужен также синтез актомиозинового комплекса. Процесс секреции подразумевает фазу собственно секреции и фазу синтеза ( накопления) секрета. Нервная ткань функционирует по такому же принципу, так как возбуждение мембраны нейрона заканчивается в конечном итоге секрецией - выбросом медиатора в синаптическую щель (изменение ЭЭГ и скорости проведения импульса в клинической практике позволяет опосредованно оценить способность нейрона к количественному синтезу медиатора). В соединительной ткани происходят те же процессы, только ее функция отличается возможностью синтезировать коллаген, эластин и пр. [17]. Исходя из вышесказанного, можно дать следующее определение функции: "Функция - это переменная величина, характеризующая циклический процесс синтеза ( накопления) и распада ( выделения) специфического органического субстрата." В соответствии с математическим определением функции специфический органический субстрат является аргументом данной функции и именно от его изменения зависит переменная величина функции. Лучшему пониманию изложенного материала поможет таблица. Обратите внимание, что в ней различны только специфический субстрат и специфическая функция, остальные пункты едины для всех клеток и тканей. Таким образом, когда речь идет о функции, то подразумеваются две фазы: фаза проявления специфической функции и фаза возврата к исходному уровню. Примером графического изображения функции являются ЭКГ, электромиограмма, доплерография. Однократное проявление специфической функции клетки, т.е. однократное сокращение, однократное выделение секрета - это еще не признак жизнеспособности и жизнедеятельности [1, 4, 5, 13]. Например, у трупа можно вызвать одиночные сокращения мышечных групп и внутренних органов после воздействия на них электрическим разрядом, но это не значит, что ткань (орган) функционирует. Значит, жизнеспособность - это способность к многократному периодическому проявлению своей специфической функции. А жизнедеятельность - это многократное проявление специфической функции. Процессы, происходящие при проявлении функции
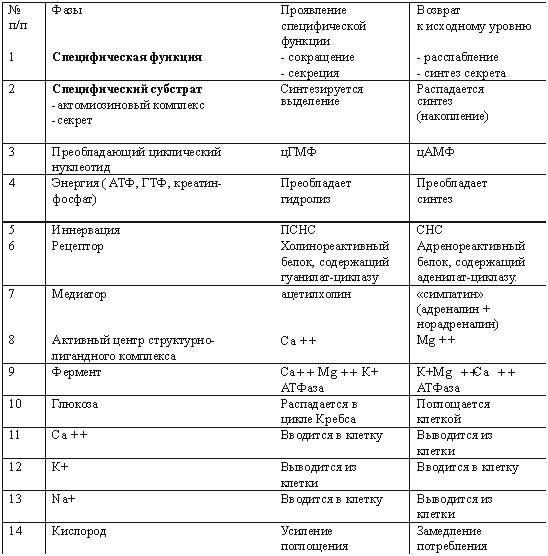
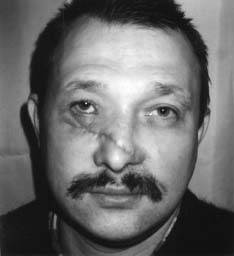
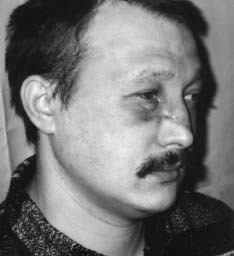
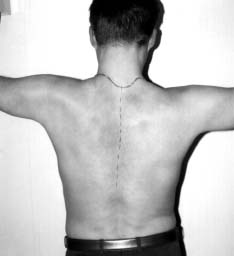
Например, ритмические сокращения сердечной мышцы у изолированного сердца свидетельствуют о высокой жизнеспособности этого органа и его возможности проявлять жизнедеятельность путем синтеза и распада большого количества актомиозиновых комплексов и выделения продуктов метаболизма. Всеобщий закон биологии Бауэра гласит: "Живая система никогда не находится в равновесии и все время совершает за счет своей свободной энергии работу против равновесия, устанавливающегося при данных внешних условиях" [10]. Процессы распада и синтеза в живой клетке идут параллельно, то есть эти процессы являются процессами с отрицательной энтропией, так как они противодействуют увеличению энтропии, связанному с распадом структур. Прекращение этого процесса означает потерю структурности, смерть. Труп переходит в состояние термодина мического равновесия, характеризующегося возрастанием энтропии [10]. В отличие от машин живые организмы используют энергию, во-первых, для совершения внутренней работы - создания и поддержания структур, т.е. синтеза белков на матрице ДНК, во-вторых, для осуществления внешней работы. Оба процесса зависимы друг от друга и протекают одновременно [10]. Процесс синтеза и распада белковых субстратов всегда идет параллельно. ДНК является носителем генетической информации и именно на ее базе формируются как структурные, так и специфические белки, определяющие специализацию клетки и ее специфическую функцию [7, 8, 10]. Согласно теории П.К. Анохина [2,15], любая функция проявляется в ответ на изменение факторов внешней среды, носит защитный (приспособительный) характер, что определяет ее целесообразность, и характеризуется процессом синтеза и распада специфического органического субстрата (см. выше определение функции). Значит, понятие "жизнь" в реабилитологии можно сформулировать так : " Жизнь - это способ передачи генетической информации во времени и защита ее в пространстве белками, синтезиру емыми на базе этой генетической информации". Это определение не противоречит ни одному из ранее данных определений жизни Н.П. Дубинина, К. Перре, Н.Т. Костюка [12]. Помогает реабилитологам понять, что в ответ на любой фактор внешней среды (а реабилитация - это совокупность внешних воздействий на организм с целью восстановления сниженной функции ) живая система будет синтезировать специфические белки, если ее генетический аппарат сохранен на клеточном уровне. Об этом же свидетельствует принцип Ле-Шателье в биохимии и биофизике [3, 6, 17]. Например, занятия ЛФК приводят к восстановлению дистрофически измененной мускулатуры в ответ на изменение градиента нагрузок (давления) на живой организм. В спорте градиент нагрузок заставляет генетический аппарат спортсменов усиленно синтезировать актин и миозин, чем и объясняется увеличение объема мускулатуры. Данное выше определение функции позволяет не разграничивать понятия "функциональные" и "органические" изменения при патологических процессах, а принимать их лишь как различные стадии дистрофического процесса [1, 7, 8]. Мы можем говорить о необратимости этих процессов только в случае, если поражен генетический аппарат клетки (пикнолиз) или всей ткани (некроз), и эта клетка, ткань, орган не могут синтезировать белки, т.е. проявлять свою специфическую функцию. В случае сохранения генетического аппарата всегда имеется возможность восстановления функции этой ткани и органа. Исходя из вышесказанного, можно дать определение понятию необратимого состояния - биологической смерти. "Биологическая смерть - это всегда деструктуризация генетического аппарата", а не внутриклеточных структур. Если нет повреждения ядра, то нужно говорить о снижении функции или ее видоизменении, а не о потере жизнеспособности, т.е. о дистрофии, состоянии обратимом, как бы далеко этот процесс ни зашел. Например, некоторые узники концлагерей (у которых все органы и ткани были в состоянии крайней дистрофии, но при нормальном уходе, питании и реабилитации были восстановлены) живы до сих пор и проявляют активную жизнедеятель ность. Для восстановления функции у них было сохранено главное - генетический аппарат в клетках тканей, который постепенно стал производить специфические белки в ответ на изменившиеся факторы внешней среды. Возможность выхода из стадии с крайне выраженной нарушенной функцией подтверждают и нижеприведенные примеры из практики автора. Пример 1. У пациента К., 40 лет, в результате травмы циркулярной пилой были повреждены до надкостницы мягкие ткани лица. В результате развился большой деформирующий рубец, произошел частичный паралич мускулатуры правой половины лица, нарушилась возможность зажмуривать правый глаз. На месте специфической ткани эпидермиса, дермы, мимической мускулатуры развилась соединительная ткань, составляющая основу рубца, т.е. видоизменилась специфическая функция этих тканей (рис. 1, А). В отделении хирургической косметологии в помощи было отказано из-за наличия противопоказаний. Через год после травмы пациент обратился к нам, и ему неинструментальными методами был ликвидирован рубец (рис. 1, Б). У пациента были восстановлены симметричная мимика, чувствительность и возможность зажмуривать правый глаз. Данный пример свидетельствует о возможности трансформации соединительной ткани рубца в специфическую ткань кожи, мышц, сосудов и нервов. У клеток соединительной ткани ядро имеет генетический аппарат (23 пары хромосом), одинаковый для всех соматических клеток. При воздействии факторов внешней среды (реабилитационных мероприятий) в соответствии с законом местоположения генетический аппарат клеток стал синтезировать те специфические белки, которые соответствовали тканям, поврежденным циркулярной пилой, т.е. реабилитационные мероприятия привели к регенерации тканей. Значит, регенерация тканей - это проявление тканями возможности просинтезировать специфический белок, который и определяет специфическую функцию этих тканей. Восстановление функции опорно-двигательного аппарата - это процесс, который характеризует возможность восстановления специфических функций большого количества тканей, окружающих суставы при патологических процессах, как бы далеко они ни зашли. Пример 2. Пациент Д., 33 лет, при спуске с горы на лыжах упал на левое плечо. Падение привело к отрыву большого бугорка плечевой кости. У пациента развился посттравматический плечелопаточный периартрит (нарушились приведение и отведение плеча (рис. 2, А). После безуспешных попыток восстановления движений в плечевом суставе реабилитологами Канады через год после травмы пациент обратился к нам. Используя разработанные нами способы скоростной реабилитации в течение трех недель мы восстановили ему движение в плечевом суставе практически в полном объеме (рис. 2, Б), несмотря на далеко зашедший дистрофический процесс в тканях плечевого сустава и пессимистические прогнозы других реабилитологов. В контексте данной статьи этот пример говорит о том, что правильный подбор факторов внешней среды - реабилитационных мероприятий, воздействующих на организм, заставит клетки тканей переориентировать свои биохимические процессы ( в соответствии с принципом Ле-Шателье) так, что в ответ на эти факторы они начнут синтезировать специфические белки и другие органические субстраты. А это и есть не что иное, как восстановление функции (см. определение). Этот пример подтверждает, что сохранение генетического аппарата клеток позволяет восстанавливать синтез специфического белка - специфическую функцию. Неинструментальные способы реабилитации: Рис. 1. Пример регенерации тканей в области деформирующего рубца А- до реабилитации (через год после травмы)
Б - после трех месяцев реабилитации Рис. 2. Посттравматический плечелопаточный периартрит годовой давности А - до реабилитации Б -после трех недель реабилитации Выводы Четкое понимание диалектического единства структуры и функции, а также понятий "жизнь", "жизнеспособность", "жизнедеятельность", "регенерация", "смерть" позволит реабилитологам избежать ненужных споров и пессимистических прогнозов по вопросу об обратимости и необратимости патологических состояний и восстановлению нарушенных функций. Только компетентность и навыки специалис та определяют возможность изменения направления патологических процессов и их конечный результат. Понимание физиологических процессов и законов, на которых эти процессы базируются, поможет не только реабилитологам, но и спортсменам достигать быстрых положительных результатов в восстановлении утраченных функций с наименьшими трудозатратами. Необходимость четкого определения базисных понятий в реабилитологии остроактуальна, и авторы статьи предполагают вернуться к этой теме. Литература 1. Адо А.Д. Патологическая физиология. - М.: Медицина, 1980. 2. Анохин П.К. Узловые вопросы современной физиологии. - М.: НИИ им. П.К.Анохина, 1976. 3. Артюхов Б.Г., Т.А. Ковалева, В.П. Шмелев. Биофизика. Воронеж, 1994. 4. Бабский Е.Б. с соавт. Физиология человека. - М.: Медицина, 1966. 5. Вилли К., В. Детье. Биология /Пер. с англ. - М.: Мир, 1978. 6. Владимиров Ю.А. с соавт. Биофизика. - М.: Медицина, 1983. 7. Ивановская Т.В., А.В. Цинзерлинг . Патологическая анатомия. - М.: Медицина, 1971. 8. Коган Э.М., Г.Е. Островерхов . Нервные дистрофии легких. - М.: Медицина, 1971. 9. Ленинджер А. Биохимия /Пер. с англ. - М.: Мир, 1974. 10. Либберт Э. Основы общей биологии /Пер. с нем. - М.: Мир, 1982. 11. Мецлер Д. Биохимия, тт. I - III /Пер. с англ. - М.: Мир, 1980. 12. Саркисов Д.С., М.А. Пальцев, М.К. Хитров. Общая патология человека. - М.:Медицина, 1995. 13. Стерки П. Основы физиологии. - М.: Мир. 1984. 14. Стайер Л. Биохимия, тт. I - III /Пер. с англ. - М.: Мир. 1984. 15. Судаков К.В. Теория функциональных систем. М., 1996. 16. Терновой К.С. Неотложные состояния (атлас). - Киев: Здоров'я, 1984. 17. Уайт А. Основы биохимии. - М.: Мир, 1984. 18. Цибуляк Т.Н. Реаниматология. - Киев: Здоров'я, 1976. 19. Ясуо Кагава. Биомембраны. - М.: Высшая школа, 1985. Поступила в редакцию 20.06.98
При любом использовании данного материала ссылка на журнал обязательна! |