|
Abstract EVALUATION OF ADAPTIVE REACTIONS' EFFECTIVENESS AT CYCLIC MUSCULAR WORK A.S. Radchenko, doctor of pedagogy St.-Petersburg P.F. Lesgaft academy of physical culture V.E. Borilkevich, professor A.I. Zorin, doctor of pedagogy St.-Petersbutg state university, St.-Petersburg Key words: long distance runners, adaptation reactions' effectiveness, cardiorespiratory system, classification of reaction types. The purpose of the study was to elaborate the methods of the cardio-respiratory system's adaptative effectiveness in "endurance" running. The subjects (long and middle distances runners and orienteers) distributed in two experimental groups (n1 = 62; n2 = 47) had been tested many times with a help of tredmill running with anaerobic threshold's VE, HR, VO2, VE/VO2, ExcCO2 and O2-П detecting. As a result of the research it had been shown: 1) the value of VO2max, speed of running and ExcCO2 to VO2max are directly depended on the effectiveness of exercising with submaximal intensity; VO2 and above indicies demonstrate the significant diurnal fluctuation in one and the same persons; the change of the HR working intensity curve's bending moment correlates with the value of ExcCo2 when working in submaximal regime and its dynamics to VO2max.
|

ОЦЕНКА ЭФФЕКТИВНОСТИ АДАПТИВНОЙ РЕАКЦИИ ПРИ ЦИКЛИЧЕСКОЙ МЫШЕЧНОЙ РАБОТЕ А.С. Радченко, В.Е. Борилкевич,
А.И. Зорин Ключевые слова: бегуны на длинные дистации, кардиореспираторная система, адаптивная реакция, изменение градиентов, классификация типов реакции. Одной из составляющих успеха в беге на средние и длинные дистанции является величина VO2max. Значительно большее влияние на результаты в беге оказывают факторы, определяющие реализацию этого показателя [11, 12]. Взаимодействие всех звеньев систем транспорта и утилизации кислорода должно обеспечивать достижение VO2max при работе большей интенсивности. За критерий оценки взаимодействия этих систем можно взять их кислород-транспортную эффективность, которая будет оцениваться как результат взаимодействия внешнего дыхания, систолического выброса, периферического кровотока и окислительного фосфорилирования (Константинов и др., 1986). Увеличение потребления кислорода в объеме всего организма тесно связано с кислородной стоимостью усиления насосной функции сердца. Эту взаимосвязь необходимо учитывать при оценке эффективности реакции на физическую нагрузку [24]. Имеются исследования, в которых показано, что воздействие интенсивных и длительных физических нагрузок на организм спортсмена вызывает многочасовые изменения количественных характеристик системы красной крови, интенсивности тканевого дыхания и др. [2]. Изменения значений многих фоновых (предрабочих) показателей определяют большие различия в результатах последующего тестирования [6]. Причиной этого может быть изменчивость взаимодействия центральных и периферических механизмов доставки и утилизации кислорода в организме спортсмена. Нарастание физической нагрузки от VO2 покоя до VO2max обеспечивается последовательностью рекрутирования мышечных волокон (МВ) медленного, промежуточного и быстрого типов [43]. Известный феномен анаэробного порога (АнП) проявляется как результат превалирования внутримышечного накопления лактата и выхода его в кровь, с одной стороны, и активности механизмов, сдерживающих метаболический ацидоз (буферирование, утилизация лактата), - с другой [9]. Феномен АнП также проявляется в скачкообразном изменении ряда показателей внешнего дыхания [45]. Несмотря на то что "вентиляционный" АнП и "лактатный" АнП имеют высокую корреляцию [13, 9], предварительное гликогеновое истощение мышц меняет проявление "лактатного" АнП относительно "вентиляционного" АнП. Изменяются легочная вентиляция, выделение двуокиси углерода, концентрация глюкозы и лактата крови при работе, а также динамика многих показателей при работе и последующем восстановлении [38, 23, 17 и др.]. Можно предполагать, что изменения градиентов ЧСС, VO2, ExcCO2, после "вентиляционного" АнП, относительно их градиентов на нагрузках до этого порога (как более стабильного по проявлению на конкретной интенсивности работы при измененной энергетике мышц) будут являться характеристикой эффективности адаптивной реакции организма спортсмена на физическую нагрузку. Мерой эффективности при этом будет количество преодолеваемой дистанции, приходящейся на единицу потребления кислорода при стандартной ЧСС. Целью настоящего исследования является разработка критериев оценки эффективности адаптивной реакции на физическую нагрузку с применением распространенных способов анализа внешнего дыхания и деятельности сердца. Методы. В исследовании приняли участие две группы бегунов разного уровня подготовленности. В первую группу были включены бегуны на средние (имевшие результаты на 1500 м от 3.00 до 4.00 с), длинные дистанции (5000 м - 13.20-15.00), марафонцы (2:13.50 - 2:28.0). Вторую группу составили кроссмены-ориентировщики, имевшие более низкие результаты в беге на длинные дистанции. Каждый испытуемый проходил предварительное тестирование с целью определения АнП по данным газоанализа и легочной вентиляции. Процедура тестирования заключалась в выполнении испытуемым ступенчато повышающейся нагрузки на тредмилле. Длительность бега на каждой ступени - 3 мин, интервал отдыха - 1 мин. Дискретность увеличения скорости бега - 0,1 м с-1. В ходе эксперимента регистрировали скорость бега V (мЧс-1), ЧСС (удЧмин-1), легочную вентиляцию - Ve (лЧмин-1), процент содержания кислорода и углекислого газа в выдыхаемом воздухе - %O2, %CO2, потребление кислорода VO2 (млЧкг-1Чмин-1), излишки углекислого газа - ExcCO2 (млЧкг-1 мин-1), кислородный пульс - О2 - П (млЧуд-1Чкг-1), вентиляционный эквивалент по кислороду - VE/VO2. АнП определяли по характерному скачку в динамике Ve, Ve/VO2 и других показателей [7, 3, 13]. Газоанализ проводился радиочастотным масспектрометром МХ6202. ЧСС и легочная вентиляция измерялись прибором "Физиолог-3" с использованием датчика дыхания турбинного типа производства "Биофизприбор". Расположение электродов для регистрации ЭКГ - по Нэбу.
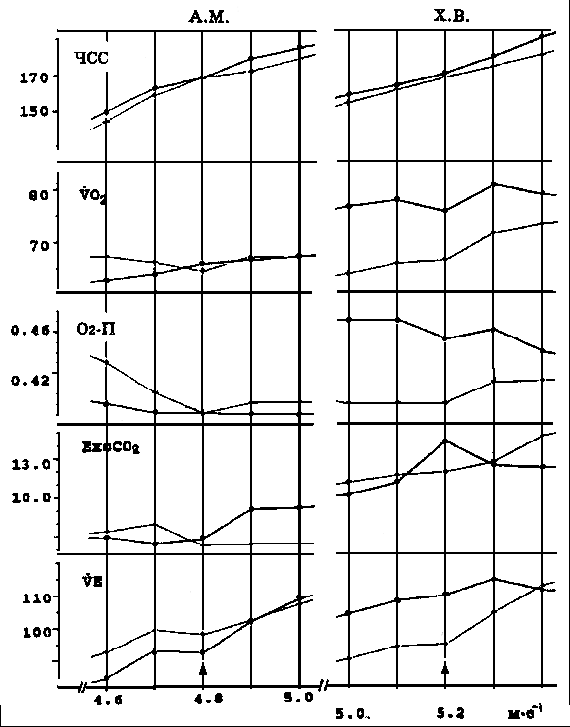
Рис. 1. Результаты повторного тестирования, испытуемый А.М. (1500 - 3.58,6) и Х.В. (марафон - 2.17,08). По абсциссе - скорость бега на тредмилле, по ординате - данные HR (B*min-1), VO2(ml*kg-1*min-1), O2P(ml*kg-1*b-1), ExcCO2(ml*kg-1*min-1), VE(l*min-1). Линии с точками - данные 1-го дня, линии с плюсами - данные 2-го дня. Стрелка - скорость бега, соответствующая АнП. В 1-й серии экспериментов приняли участие 18 бегунов из 1-й группы. Изучалась динамика состояния каждого из них после предварительной интенсивной и длительной тренировочной нагрузки - равномерного бега на уровне индивидуального АнП в течение 32-34 мин (10 км) на стадионе. На следующий день и далее 3-4 дня подряд в одно и то же время суток проводились тестирования этого спортсмена на тредмилле с целью определения АнП. Диапазон нагрузок выбирался с тем расчетом, чтобы ступень предполагаемого АнП была средней по порядку предъявления. Вычисляли коэффициент регрессии для значений показателей, полученных на последней минуте каждой ступени нагрузки до АнП и симметрично для значений тех же показателей, полученных на ступенях нагрузки выше АнП (3-4 ступени до АнП и столько же после АнП). Сопоставление градиентов перечисленных показателей до АнП и после его превышения составляет основу последующего анализа. Результаты такого подхода к оценке адаптивной реакции сопоставлялись с традиционными количественными показателями. Так, эффективность работы оценивалась по кислородной стоимости выполняемого упражнения на субмаксимальной интенсивности (при беге с ЧСС=170 уд/мин-1). Вычислялось количество метров преодолеваемой дистанции, приходящихся на 1 млЧкг-1 потребляемого кислорода - показатель Q (мЧкгЧмл-1). В тех случаях, когда при тестировании удавалось достигнуть VO2max, для оценки эффективности адаптивной реакции в расчет бралась его величина, а также ЧСС, ЕхсСО2 и скорость бега при этом. Все статистические вычисления проводились с использованием стандартной компьютерной программы. Во 2-й серии экспериментов принимали участие спортсмены обеих групп. Их состояние оценивали в течение одного дня без применения предварительной интенсивной и длительной тренировочной нагрузки. После 2-3 дней легких тренировок выполняли тестирование для определения АнП. Значительная часть испытуемых обследовалась многократно с интервалом от 2-3 недель до 2-3 месяцев. В экспериментах приняли участие 60 бегунов и 47 ориентировщиков. Средние данные испытуемых приведены в табл. 1. Таблица 1. Средние данные испытуемых (mean ± SE)
Таблица 2. Результаты тестирования одних и тех же испытуемых два дня подряд (tgY)
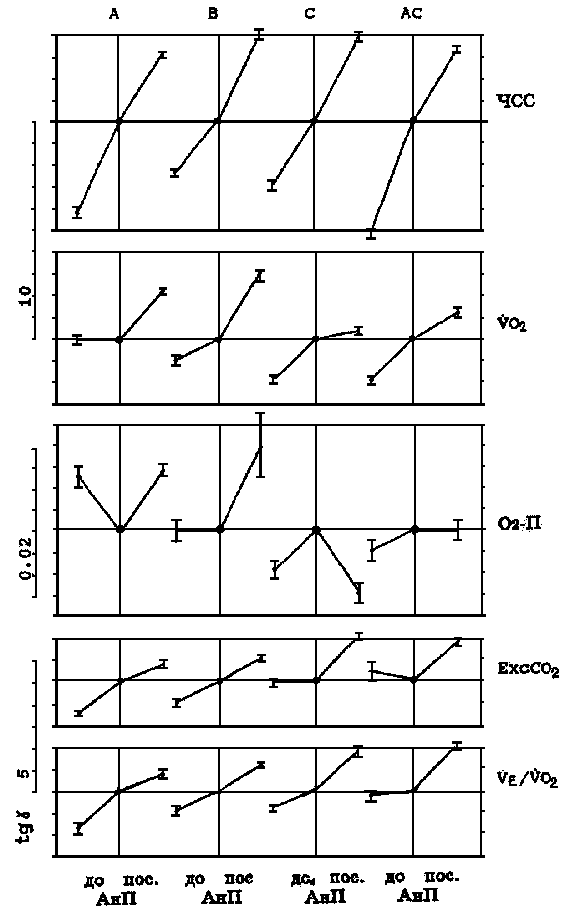
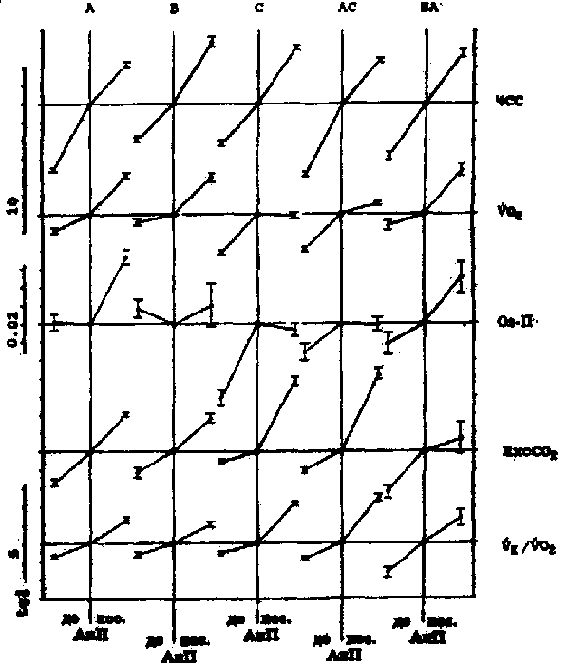
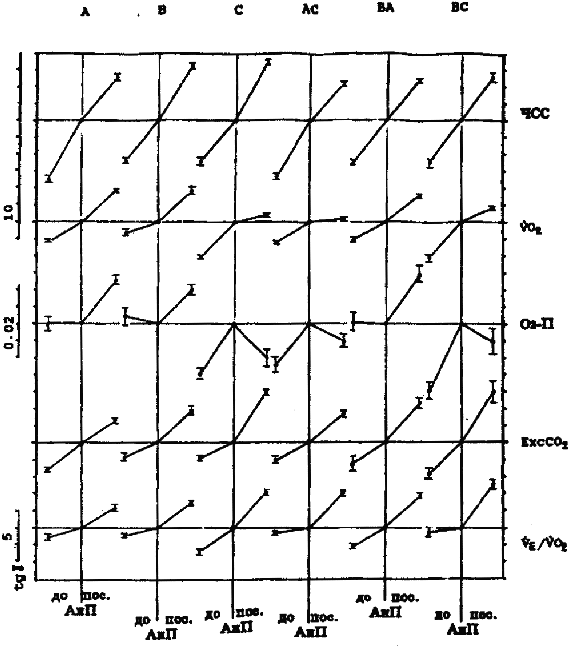
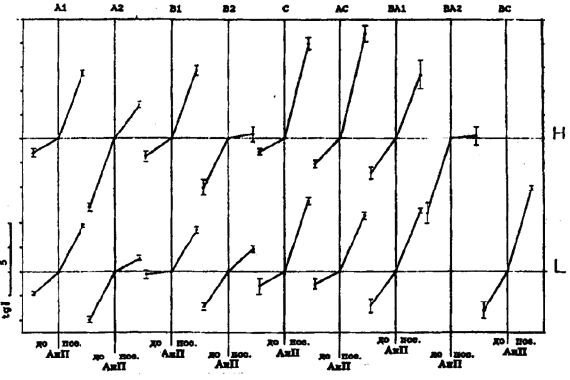
Примечание. Q - эффективность выполнения тестирующей нагрузки (количество метровпреодолеваемой дистанции, приходящейся на 1 мл*кг-1, потребляемого кислорода при беге с ЧСС = 170 уд/мин-1). Результаты. Обратим внимание на то, что кардиореспираторные показатели, зарегистрированные на одних и тех же ступенях нагрузки у одного и того же спортсмена, значительно отличаются по дням (рис. 1). Существенно различаются и градиенты этих показателей при увеличении нагрузок в зоне аэробно-анаэробного перехода и АнП (табл. 2). Если рассматривать данные на рис. 1 с позиций теста PWC170, то показатели и первого, и второго дня у обоих испытуемых равнозначны (см. рис. 1). ЧСС в 170 удЧмин-1 у первого испытуемого соответствует скорости бега между 4,8-4,9 мЧс-1, а у второго - 5,2-5,3 мЧс-1. В то же время очевидно, что динамика кардиореспираторных показателей разная и поддержание задаваемой нагрузки обеспечивается, вероятно, различной комбинацией взаимодействия систем транспорта и утилизации кислорода. Итак, результаты, полученные в 1-й серии экспериментов, показали, что у одних и тех же испытуемых темп нарастания ЧСС после превышения нагрузки АнП может увеличиваться или уменьшаться. То же самое происходит и с VO2. Независимо от ЧСС градиент VO2 может или увеличиваться или уменьшаться. Далее классифицируем встретившиеся типы реакции, взяв за основу направленность динамики изучаемых показателей. Средние данные этой серии представлены на рис. 2. Так, градиент ЧСС с превышением нагрузки АнП в одних случаях уменьшается (реакции типа А, АС), а в других - увеличивается (реакция типа В, С). Градиент VO2 с превышением нагрузки АнП в одних случаях увеличивается (реакции типа А, В), а в других - уменьшается (реакции типа С, АС). Соответственно меняется и динамика О2 - П. Результаты, полученные во 2-й серии экспериментов, были обработаны по такому же принципу (рис. 3 и 4). В них обнаружилось еще два типа реакций - ВА, ВС. Градиент ЧСС не меняется во всем диапазоне нагрузок. Градиент VO2 в первом случае увеличивается (реакция ВА), в другом - уменьшается (реакция ВС). Обнаружилось также, что в одних и тех же типах реакций (А, В, ВА) в одних случаях наблюдается значительное увеличение, а в других - значительное уменьшение градиента ЕхсСО2. Случаи с уменьшением градиента ЕхсСО2 выделим как отдельный тип реакции и обозначим далее как реакции А2, В2 и ВА2 (рис. 5). Уменьшение градиента ЕхсСО2 составило до 26,7% от общего числа результатов тестирований во 2-й серии у 1-й группы и 22,8% у 2-й.
Рис. 2. Результаты 1-й серии экпериментов (M ± m). По ординате - абсолютная величина коэффициэнта регрессии (цена шкалы для HR, VO2, O2P и т.д. указана слева). A, B, C, AC - типы реакций.
Рис. 3. Результаты 2-й серии экпериментов - 1- группа испытуемых (бегуны высокого класса) - M ± m. Обозначения те же, что и на рис. 2.
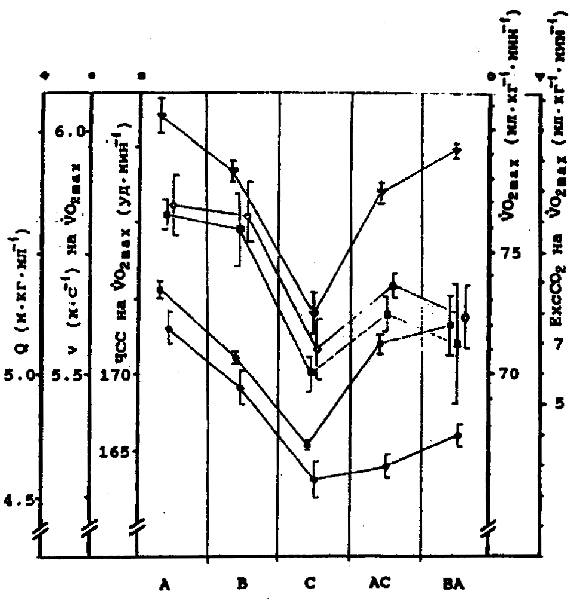
Рис. 4. Результаты 2-й серии экпериментов - 2- группа испытуемых (ориентировщики) - M ± m. Обозначения те же, что и на рис. 2. Сравнение различных типов реакций по эффективности выполнения упражнения показало, что у испытуемых 1-й группы в реакциях типа А эффективность выполнения упражнения выше, чем в других типах реакций (см. рис. 6, показатель Q). В реакциях типа С у тех же лиц эффективность ниже. Эти данные были получены при многократном тестировании одних и тех же лиц. На основе t-критерия Стьюдента для связанных выборок различия достоверны между данными реакций А и В, А и С, А и АС, В и АС (p < 0,01) у испытуемых 1-й группы. Здесь же представлены данные, полученные при тестированиях, когда удалось достигнуть VO2max. Так, у испытуемых 1-й группы при разном типе реакций скорость бега на VO2max значительно меняется. В реакциях типа А она выше, чем в реакциях типа В, С, АС и ВА. В реакциях типа С эта скорость самая низкая по сравнению со всеми типами реакций. Более высокой скорости бега на VO2max соответствуют более высокие ЧСС и ЕхсСО2 у одних и тех же бегунов в разные дни.
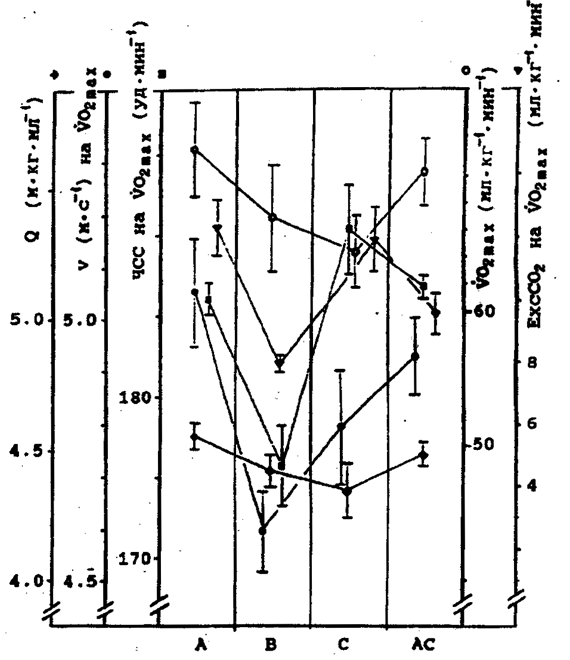
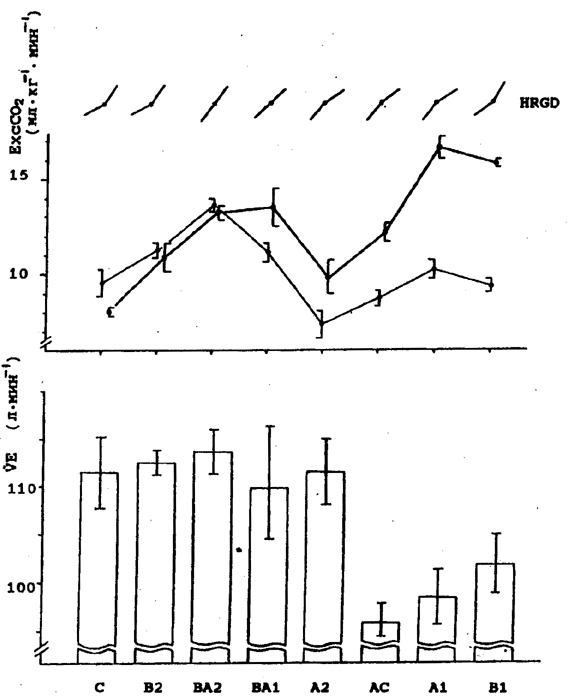
Рис. 5. Динамика ExcCO2 у 1-й (H) и 2-й (L) группы испытуемых по результатам 2-й серии экспериментов. Обозначения те же, что и на рис. 2. У испытуемых 2-й группы в результатах обнаруживается схожа тенденция только в эффективности реакций и величине VO2max. В других показателях подобных закономерностей не обнаруживается (рис. 7). Далее на абсциссе расположим имеющиеся типы реакций по мере увеличения разницы между ЕхсСО2 при беге на субмаксимальной интенсивности и ЕхсСО2 на VO2max (рис. 8). Видим, что постепенному увеличению - ЕхсСО2 соответствует последовательное изменение динамики ЧСС (HRGD) - верхн часть рис. 8. Нетрудно заметить также, что легочная вентиляция намного больше при 95% VO2max в тех случаях, когда величина -ЕхсСО2 небольшая или отрицательная. VE резко снижена, когда наблюдается увеличение - ЕхсСО2 - нижняя часть рис. 5. Во 2-й группе испытуемых количество случаев соответствующих реакций оказалось недостаточным. Отметим, что в 1-й группе - бегуны высокого класса - АнП проявлялся на скорости бега (mean ± SE) 5,26±0,08 мЧс-1, во 2-й - ориентировщики - на скорости бега 4,43±0,12 мЧс-1. Обсуждение. Известно, что эффективность насосной функции сердца может характеризоваться направленностью градиента ЧСС после АнП относительно его величины до АнП. Так, уменьшение градиента ЧСС после АнП говорит о большей эффективности работы сердечного насоса, поскольку в этом случае нарастание кровотока обеспечивается большей фракцией изгнания левого желудочка (LVEF), особенно на пике нагрузки [20, 33]. Нами показано, что кривая ЧСС / интенсивность работы в зоне АнП далеко не всегда имеет изгиб, как считалось ранее [10, 42]. Более того, обнаруживается не только сглаживание точки изгиба кривой при интенсивности работы соответствующей АнП, но и обратное явление - увеличение градиента ЧСС после АнП, на что указывают многие авторы [16, 20, 33].В нашем исследовании у испытуемых 1-й группы в 7,4% случаев изгиб не обнаруживается, а в 34,3% случаев градиент ЧСС увеличивается. Во 2-й группе испытуемых - 6,8 и 30,4% соответственно. По результатам исследований [20] аналогичная динамика ЧСС наблюдается в 3,3 и 5,6% случаев соответственно. Заметное отличие от наших данных объясняется принципиально иным протоколом тестирования, а также отличиями в контингенте испытуемых и средствах предъявления нагрузки. Кроме того, мы ориентируемся на "вентиляционный порог", а названные авторы - на "точку излома кривой лактата". Учитывая наши данные, теперь можно сказать, что систолический выброс при нарастании физической нагрузки обеспечивается различной комбинацией ЧСС и ударного объема у одного и того же человека в разные дни. К этому добавим, что потребление кислорода миокардом непосредственно зависит от величины постнагрузки сердца [36]. Напряжение стенок желудочка, а также скорость его сокращения - факторы, прямо определяющие потребление кислорода миокардом [39, 34]. Следовательно, увеличение градиента ЧСС после превышения интенсивности упражнения соответствующей АнП характеризует усиление влияния этих факторов. Таким образом, реакции А и АС могут быть более эффективными по отношению к реакциям В и С не только по характеристике производительности сердца, но и по критерию энерготрат миокарда. Реакции ВА и ВС по динамике ЧСС занимают промежуточное положение между реакциями типа А, В, С. Вероятность проявления точки изгиба на кривой ЧСС/интенсивность работы при превышении физической нагрузки, соответствующей резкому увеличению накопления лактата в крови, может быть объяснена взаимодействием разных механизмов рефлекторного контроля кровотока. С предъявлением физической нагрузки импеданс артериальной системы значительно снижается - примерно в три раза [4]. В покое и при работе умеренной интенсивности (40% от VO2max) раскрытие сосудистого русла обеспечивается барорефлекторным контролем [14, 19, 31, 40]. Когда интенсивность упражнения значительно возрастает и концентрация метаболитов (лактат, Н+) в работающих мышцах увеличивается, активируются хемoсенсорные афференты [32, 44]. При выполнении упражнений максимальной интенсивности и максимальной производительности сердца, когда общая проводимость сосудов в работающих мышцах достигает 70% и более и как только заполнится сосудистое русло, поддержание давления крови мышечными хеморефлексами может потребовать сужения просвета сосудов вовлеченных в работу мышц [35, 41, в публикации]. Данные на рис. 8 хорошо демонстрируют эту концепцию. Можно с уверенностью предполагать, что различное состояние углеводного резерва мышц, которое меняется у повседневно тренирующегося спортсмена, определяет динамику хеморефлекторной регуляции после превышения нагрузки АнП. Особенно отчетливо это проявляется, как видим, не просто у хорошо тренированных спортсменов, а у специально подготовленных спортсменов высокого класса при адекватных условиях тестирования.
Рис. 6. Данные, полученные при повторных тестированиях одних и тех же лиц. 1-я группа испытуемых (бегуны высокого класса) - M ± m, n=21. По абсциссе -типы реакций Когда интенсивность физической нагрузки начинает превышать АнП, рекрутируются МВ быстрого типа, а волокна медленного типа уже полностью вовлечены в работу [7, 9]. Следует особо подчеркнуть, что и рекрутирование МВ промежуточного типа продолжается до нагрузки по крайней мере в 92% от VO2max [43]. Если учитывать метаболические характеристики МВ [21], то это означает, что доля энергопродукции в мышцах за счет активности аэробного гликолиза может значительно увеличиваться. Скорость выработки энергии при этом будет выше примерно наполовину [30] по сравнению с использованием окисления жирных кислот. Такая ситуация, возможно, возникает в реакциях типа А, В, ВА, и увеличение градиента VO2 обусловлено названными причинами. Уменьшение градиента VO2 в реакциях типа С, АС, ВС может происходить, если нарастание физической нагрузки после АнП обеспечивается использованием жирных кислот и рекрутированием МВ быстрого типа. Доля участия МВ с аэробным гликолизом в поддержании работы выше интенсивности АнП уменьшается по причине изначально сниженного углеводного резерва. Такое объяснение может быть принято, поскольку изменение параметров "лактатного" АнП - интенсивности рабочей нагрузки, %VO2max, концентрации лактата крови - относительно параметров "вентиляционного" АнП отчетливо наблюдается у предварительно гликоген-истощенных субъектов [23 и др.]. В реакциях, где превышение нагрузки соответствующей АнП сопровождается уменьшением градиента ЕхсСО2 (реакции А2, В2, ВА2), обнаруживается значительно повышенная на (95%VO2max) легочная вентиляция у одних и тех же испытуемых (см. рис. 8). По аналогии с данными Segal и Brooks (1979) можно предполагать, что гликогеновый резерв при этих типах реакций также изначально снижен, однако увеличение градиента VO2 после АнП все же обнаруживается. Убедительное объяснение может быть дано после выявления степени активности механизмов глюконеогенеза печени и коры почек, который использует лактат и некоторые недоокисленные продукты и аминокислоты при интенсивной мышечной работе. Заметим, что суммарное количество глюкозы, выходящее при этом в кровь, может покрывать до половины максимальной потребности мышц в ней [1, 8, 15, 18, 27], а фармакологическое стимулирование глюконеогенеза дает хороший эффект повышения работоспособности и купирования утомления [1]. Становится очевидным тот факт, что при каждом тестировании проявляется иная стратегия адаптации в мышцах с преимущественной мобилизацией того или иного субстрата для производства АТФ с изменяющейся эффективностью выполнения упражнения и различной интенсивностью использования кислорода. В разные дни у одного и того же спортсмена нарастание физической нагрузки может обеспечиваться, вероятно, изменяющейся долей участия МВ различного типа в зоне нагрузок, соответствующих аэробно-анаэробному переходу и превышающих АнП.
Рис. 7. Данные, полученные при повторных тестированиях одних и тех же лиц. 2-я группа испытуемых (ориентировщики), n=11. Обозначения те же, что и на рис. 6. Следует добавить, что поток кислорода в тканях, начинаясь в мелких и мельчайших артериолах и продолжаясь в капиллярах, частично возвращается в венулы и тем самым образует поле напряжений кислорода. Существование поля напряжений кислорода является основой для развития компенсаторных реакций [24-26]. Все механизмы его регулирования не допускают повышения парциального давления в большой массе тканей, сохраняя его на относительно низком уровне. Учитывая раскрытие резервных капилляров в мышцах при предъявлении физической нагрузки [22, 28 и др.], особенности регуляции скоростей кровотока в микрососудах, можно утверждать, что диффузи кислорода в тканях - процесс инертный и не подвержен резким изменениям. Скорость диффузии вполне соответствует необходимой скорости (объему) окислительного фосфорилирования в клетке, а увеличение капилляризации адаптированных к повышенной физической нагрузке мышц не влечет за собой увеличения периферического сопротивления кровотоку [24-26]. Таким образом, данные, приведенные на рис. 6 и 8, демонстрируют результат взаимодействия систем транспорта и утилизации кислорода. Очевидно, что именно взаимодействие названных систем во многом определяет эффективность работы сердца. Это обусловливает ежедневные изменения величины критической скорости у бегунов высокого класса с соответствующими изменениями величин всех других показателей.
Рис. 8. Верхн часть. Изменение dExcCO2 между нагрузкой при ЧСС=170 уд*мин-1 (линии с черной точкой) и нагрузкой на VO2max (линии со светлой точкой) и изменения градиента ЧСС (HRGD) до АнП и после его превышения. Нижняя часть. Величина VE при 95% VO2max. По абсциссе - типы реакций по мере увеличения dExcCO2 Основной механизм, объединяющий системы транспорта и утилизации кислорода, обнаруживается во взаимозависимости объемной скорости коронарного кровотока и парциального давления кислорода в миокарде, с одной стороны, и насыщения кислородом и его парциального давления в аортальной крови - с другой [37]. Используя данную закономерность для наших рассуждений, можно сделать хоть и приблизительный, но важный вывод: увеличение потребления кислорода на 50% в масштабе всего организма требует увеличения обеспечения миокарда кислородом на 150-200% [24]. Если взять эту взаимозависимость за основу для оценки эффективности реакции организма на физическую нагрузку, можно утверждать, что несколько иная стратегия включения метаболических путей в работающих мышцах [по 19] способна значительно изменить как потребление кислорода миокардом, так и регуляцию сердечной деятельности, а следовательно, поменять интенсивность работы, при которой достигается VO2max. Продемонстрированные нами результаты и предложенные объяснения хорошо укладываются в рамки этих положений. Заключение. Анализ динамики кардиореспираторных показателей выявляет значительные колебания эффективности реакций на циклическую физическую нагрузку у одних и тех же спортсменов. Несмотря на то что рассматриваемые показатели являются результатом многоуровневого взаимодействия различных физиологических механизмов, долю участия которых невозможно оценить отдельно друг от друга, однозначная оценка эффективности реакции на нагрузку получается посредством сопоставления динамики характеризующих их показателей. Изменчивость взаимодействия систем транспорта и утилизации кислорода у одного и того же спортсмена особенно ярко проявляется у субъектов, специально подготовленных к данному виду локомоций. Эффективность выполнения упражнения с субмаксимальной интенсивностью в целом зависит, с одной стороны, от эффективности работы сердечного насоса, его производительности и собственных энергозатрат, с другой - от стратегии адаптации энергетики мышц. Это позволяет при реакции более эффективного типа выполнять более интенсивную работу, мобилизуя значительно большие энергетические резервы. Совершенно очевидно, что одна и та же тренировочная работа, вызывая разный тип адаптивной реакции, в каждом случае будет оказывать неодинаковый тренирующий эффект. Будут отчетливо проявляться универсальные физиологические законы: ответная реакция целостного организма (любого его структурного уровня) зависит не только от интенсивности и длительности стимула, но и от состояния, в котором организм находится; стратегия адаптивной реакции - прежде всего от состояния систем, обеспечивающих долговременную адаптацию. Литература 1. Бобков Ю.Г., Виноградов В.М., Катков В.Ф. и др. Фармакологическая коррекция утомления. - М.: Медицина, 1984. 2. Евгеньева Л.Я., Бринзак В.П. Взаимосвязь систем организма при напряженной мышечной деятельности и в процессе восстановления. /Кислородные режимы организма, работоспособность, утомление при напряженной мышечной деятельности. - Вильнюс: Гос. Комитет ЛитССР по ФК, 1989, ч. 1, с. 61-67. 3. Зорин А.И. Определение анаэробного порога с помощью комплекса респираторных показателей. Вопросы физического воспитания студентов. - СПб.: Университет, 1992, т. 33, с. 109-111. 4. Карпман В.Л., Орел В.Р. Импеданс артериальной системы и сердечная деятельность. //Физиол. чел., 1985, № 4, c. 628-633. 5. Константинов Б.А., Сандриков В.А., Яковлев В.Ф. Оценка производительности и анализ поцикловой работы сердца в клинической практике. - Л.: Наука, 1986. 6. Радченко А.С., Борилкевич В.Е., Зорин А.И. Особенности оценки специальной работоспособности бегунов на длинные дистанции //Вопросы физического воспитания студентов /Отв. ред. В.Е. Борилкевич. СПб., 1992. Вып. 23, с. 79-87. 7. Селуянов В.Н., Мякинченко Е.Б., Холодняк Д.Б., Обухов С.М. Физиологические механизмы и методы определения аэробного и анаэробного порогов. "Теория и практика физической культуры", 1991, № 10, c. 10-18. 8. Смирнов А.В. Роль глюконеогенеза при физической деятельности. Успехи современной биологии. Т. 97, вып. 3, 1984, с. 399-412. 9. Brooks G.A. Anaerobic threshold: review of the concept and directions for future research. - Med. Sci. Sports and Exers., 1985, v. 17, p. 22-31. 10. Conconi F., Ferrari M., Ziglio P.G. et al. Determination of the anaerobic threshold by a noninvasive field test in runners. - Eur. J. Appl. Physiol., 1982, v. 52, p. 869-873. 11. Costill D.L. (A Scientific Approach to Distance Running) La course de fond Approche scientifique. Paris, VIGOT, 1981. 12. Costill D.L., Thomason H., Roberts E. Fractional utilization of the aerobic capacity during distance running. - Med. Sci. Sports and Exers., 1973, v. 56 p. 248-252. 13. Davis H.A. Anaerobic threshold: review of the concept and directions for future research. - Med. Sci. Sports and Exers., 1985, v. 17, p. 6-18. 14. Ebert T.J. Baroreflex responsiveness is maintained during isometric exercise in humans. J. Appl. Physiol., 1986, v. 61, p. 797-803. 15. Exton J.H. Gluconeogenesis. Metabolism, 1972, N 10, v. 21, p. 945-953. 16. Hech H., Bechers K., Lammerschmidt W. et al. Identification, оbjectivity and validity of Conconi threshold by cycle stress tests. - Dtsch. Z. Sportmed., 1989, v. 40, p. 388-412. 17. Heigenhauser G.J.F., Sutton J.R., Jones N.L. Effect of glycogen depletion on the ventilatory response to exercise. J. Appl. Physiol., 1983, v. 54, p. 470-474. 18. Hirche H., Grun D., Waller W. Utilization of carbohydrates and free fatty acids by the gastrochemius of the dog during long lasting rhythmical exercise. Pflugers Arch. ges. Physiol., 1970, B. 231, H. 2, S. 121-132. 19. Hochachka P.W., Somero G.N. Biochemical Adaptation. - Princeton, Princeton University Press, 1984. 20. Hofmann P., Pokan R., Leitner H., Schmid P. Differences in the degree and direction of the deflection of the heart rate performance curve. - Current research into sport sciences. International conference during goodwill games. St.-Petersburg, 27-30 July, 1994, p. 77. 21. Hollozy J.O., Booth F.W. Biochemical adaptations to endurance exercise in muscle. Ann. Rev. Physiol., 1976, v. 38, p. 273-291. 22. Honig C.R., Odoroff Ch.L., Frierson J.L. Capillary recruitment in exercise: rate, extend, uniformity, and relation to blood flow. - Amer. J. Physiol., 1980, v. 238, p. H31-H42. 23. Hughes E.F., Turner S.C., Brooks G.A. Effects of glycogen depletion and pedalling speed on "anaerobic threshold". J. Appl. Physiol., 1982, v. 52, p. 1598-1607. 24. Ivanov K.P. The principles of energetics in organism: Theoretical and practical aspects. V. 2. Biological oxidation and its providing with oxygen. - St.-Petersburg: Nauka 1993. 25. Ivanov K.P., Kalinina M.K., Levkovich Yu.I. Blood flow velocity in capillaries of brain and muscles and its physiological significance. - Microvasc. Res., 1981, v. 22, p. 143-155. 26. Ivanov K.P., Kalinina M.K., Levkovich Yu.I. Microcirculation velocity changes under hypoxia in brain, muscles, liver, and their physiological significance. - Microvasc. Res., 1985, v. 30, p. 10-18. 27. Krebs H.A. Renal gluconeogenesis. - Advances Enzyme Regulat., 1963, v. 1, p. 385-396. 28. Krogh A. Comparative physiology of respiratory mechanisms. Philadelphia, 1941. 29. Ludbrook J. Reflex control of blood pressure during exercise. Ann. Rev. Physiol., 1983, v. 45, p. 155-168. 30. McGilvery R.W. The use of fuels for muscular work. In: Metabolic adaptation to Prolonged Physical Exercise, ed. H.Howarld and R. Poortmans, Basel, Birkhauser Verlag, 1975, p. 12-30. 31. Milcher A., Donald D.E. Maintained ability of carotid baroreflex to regulate arterial pressure during exercise. Am. J. Physiol., 1981, N 241, P.H.838-H.849. 32. Mitchell J. H., Schmidt R.F. Cardiovascular reflex control by afferent fibers from skeletal muscle receptors. In: Handbook of The Cardiovascular System. V. 3. W.F. Humilton and P. Dow (Eds.). Bethesda, MD: American Physiological Society, 1983, p. 623-658. 33. Pokan R., Hoffman P., Preider K. et al. Correlation between inflection of heart rate/work performance curve and myocardial function in exhausting cycle ergometer exercise. - Eur. J. Appl. Physiol., 1993, v. 67, p. 385-388. 34. Ross J. Mechanisms of cardiac contraction what roles for preload, after load and inotropic state in failure // Europ. Heart J., 1983, N 4, suppl. A, p. 19-28. 35. Saltin B., Strange S. Maximal oxigen uptake: "old" and "new" arguments for a cardiovascular limitation. Med. Sc. in Sp. and Exers., 1992, v. 24, p. 30-37. 36. Sarnoff S., Nitchell B. The regulation of the performance of the heart. - Am. J. Med., 1961, v. 30, N 5, p. 747-771. 37. Schuchhardt S. Miocardial oxygen pressure. - Adv. Exp. Med. and Biol., 1985, v. 191, p. 21-35. 38. Segal S., Brooks G. Effects of glycogen depletion and work load on postexercise o consumtion and blood lactate. J. Appl. Physiol., 1979, v. 47, p. 514-521. 39. Sonnenblick E. Contractility of cardiac muscle. - Circ. Res., 1970, v. 27, p. 479-481. 40. Strange S., L.B. Rowell, N.J. Christensen B. Saltin. Cardiovascular responses to carotid sinut baroreceptor stimulation during maximal exercise with arms and legs in man. - Acta Physiol. Scand. 1990, v. 138, p. 145-153. 41. Strange S. et al., in press (1992). 42. Tiedt N., Wohlgemuth B. Die statistische Kennline der Belastungsherzfrequenz, Med. Sport., 1973, v. 13, S. 87-94. 43. Vollestad N.K., Blom P.C.S. Effect of varying exercise intensity on glycogen depletion in human muscle fibres. - Acta Physiol. Scand., 1985, v. 125, p. 395-405. 44. Victor R.G, L.A. Bertocci, S.L. Pryor, R.L. Nunnally. Sympathetic nerve discharge is coupled to muscle cell pH during exercise in humans. J. Clin. Invest., 1988, v. 82, p. 1301-1305. 45. Wasserman K., McIlroy M.B. Detecting the threshold of anaerobic metabolism. Am. J. Cardiol., 1964, v. 14, p. 844-852. 46. Wasserman K., Whipp B.J., Koyal S.N., Beaver W.L. Anaerobic Threshold and respiratory gas exchange during exercise. J. Appl. Physiol., 1973, v. 35, p. 236-243. Поступила в редакцию 18.04.96
При любом использовании данного материала ссылка на журнал обязательна! | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||