НЕЙРОГОРМОНАЛЬНОЕ ВЗАИМОДЕЙСТВИЕ В ПРОЦЕССЕ ВЫПОЛНЕНИЯ СПОРТСМЕНАМИ ТОЧНОСТНЫХ ДВИГАТЕЛЬНЫХ ДЕЙСТВИЙ
Ткачук В., Полищук Т., Ягелло В., Ягелло M.
Институт Спорта Академии физического воспитания Юзефа Пилсудского в Варшаве
В работе показана взаимосвязь между содержанием различных гормонов надпочечников в крови и моче с точностью воспроизведения спортсменами заданного пространства.Аннотация.
Ключевые слова: спортсмен, борьба, классический стиль, гормоны, надпочечники, точность, воспроизведение, движение.
Анотацiя. Нейрогормональна взаемодiя в процесi виконання спортсменами точнiсних рухових дiй. В працi показана взаемозалежнiсть помiж кiлькiстю рiзних гормонiв наднирек i точнiстю вiдтворення спортсменами заданого простору.
Ключовi слова: спортсмен, боротьба, класичний стиль, гормони, надпочечники, точнiсть, вiдтворення, рух.
Annotation. Tkachuk V., Polischuk T., Yagello V., Yagello M. Neurohormonal interplay during performance by the sportsmen accuracy of motorial operations. In work the correlation between a content of different hormonums of paranephroses in a blood and urine with a fidelity of reproduction by the sportsmen of given space is rotined.
Keywords: sportsman, wrestling, classic style, hormonums, paranephroses, accuracy, procreation, locomotion.
Краткая справка
Еще в прошлом веке стало ясно, что организме животных и человека есть система органов, выделяющих в кровь и лимфу особо активные вещества - гормоны, которые развиваются из нескольких источников и их деятельность начинает проявляться на самых ранних этапах зародышевой жизни. Эти вещества представляют собой сильнодействующие агенты. Поэтому для получения специфического эффекта достаточно даже небольшого их количества.
Анатомически обособленные железы внутренней секреции функционально оказывают влияние друг на друга. В связи с тем что это влияние обеспечивается гормонами, доставленными кровью к органам-мишеням, принят термин "гуморальная регуляция". Работают эти системы по принципу положительной и отрицательной обратной связи [1].
Для поддержания нормального роста, жизнедеятельности и адекватного развития организма необходим определенный уровень содержания гормонов в крови.
Одним из важных свойств гормонов является высокая биологическая активность, но, в тоже время, их действие довольно кратковременно.
Основные функции гормонов это:
- поддержание внутренней среды организма,
- участие в процессах метаболизма,
- регуляция роста и развития организма [Greep R.O. a. all., 1975; 2].
Эндокринная система работает во взаимодействии и под контролем ЦНС, и совместно с последней осуществляет регуляцию и координацию многочисленных функций организма. Общим, как для нервных, так и эндокринных клеток является выработка регулирующих факторов.
При помощи секреции гормонов эндокринная система, совместно с нервной, обеспечивает существование организма как единого целого. Эндокринные клетки производят гормоны и выделяют их в кровь, а клетки нервной системы (нейроны) вырабатывают биологически активные вещества (нейромедиаторы - норадреналин, ацетилхолин, серотонин и др.).
Связующим звеном между эндокринной и нервной системами служит гипоталамус, который является одновременно и нервным образованием, и эндокринной железой.
Гипоталамус представляет собой высший центр эндокринной системы. Он контролирует и объединяет эндокринные механизмы регуляции с нервными, являясь также мозговым центром вегетативной нервной системы.
Механизм действия гормонов и сравнительно прост, и, в тоже время, весьма сложен. Гормон связывается рецепторами в клетках-мишенях, при этом активируются внутриклеточные ферменты. Это приводит клетку-мишень в состояние функционального возбуждения. Поэтому слаженная работа всех органов эндокринной системы является залогом нормальной жизнедеятельности живого организма [2].
Проблемная ситуация
В сложной многоуровневой совокупности разнообразных регуляций, осуществляемых ЦНС в механизмах общей адаптации, важнейшее место занимает управление трофической функцией организма. В значительной степени эту роль ЦНС выполняет посредством регулирования функциями желез внутренней секреции (ЖВС).
Как показали исследования человека в состоянии физического и умственного напряжения отмечается не только активность различных отделов ЦНС, но и обмена веществ в нервных клетках [Митюков М.И. и соавт., 1970; Яковлев Н.Н., 1974]. В мировой физиологии эта важная связь между спортивной деятельностью и состоянием эндокринных функций была отмечена еще в 1927 году [Невядомский М.М., 1927].
Обладая большой мощностью и широким спектром действия на обмен веществ в клетках гормоны способны изменять и структуру, и функцию самих клеток, тканей и органов, т.е. их морфологию и физиологию [Розен В.Б., 1980]. А если учитывать, что в основе любой деятельности спортсмена лежит согласованная деятельность различных органов и систем организма, а в каждой их них взаимосвязанная деятельность огромного числа клеток, то эффективность выполнения функций на клеточном уровне определяется тремя основными компонентами:
- Функциональной мощностью соответствующих клеточных структур, детерминированных генетической программой и уровнем морфо - функционального усовершенствования, являющегося результатом долговременной адаптации к условиям существования, в том числе, и к уровню двигательной активности.
- Энергетическим обеспечением функции, которое заключается в совокупности энергетических процессов, в результате которых синтезируется и освобождается достаточное количество энергии для осуществления функциональной активности на требуемом уровне.
- Пластическим обеспечением функции, которое [по Ф.З. Меерсону, 1967], представляет собой процесс обновления функционирующих, энергообразующих и опорных структур клеток, осуществляемый посредством биосинтеза белка и обеспечивающий устойчивость физиологических функций в условиях целостного организма [Виру А.А., Кырге П.К., 1983].
В процессе функционирования указанных компонентов неизбежно проявление вариативности как в каждой их перечисленных функций, так и многих других. Это вытекает из того, что каждый организм представляет собой динамическое сочетание устойчивости и изменчивости, в котором изменчивость служит его приспособительным реакциям, направленным на поддержание жестких и пластичных констант организма [Анохин П.К., 1968]. Изменение функциональной активности органов и систем, выражающихся в сдвигах пластических констант организма, обеспечивает необходимые условия для выполнения актов жизнедеятельности, а также поддержание жестких констант.
В процессе функционирования перечисленных компонентов неизбежно появление феномена вариативности как в каждой из перечисленных функций, так и многих других [Ткачук В., 1986]. Это вытекает из того, что каждый организм представляет собой динамическое сочетание устойчивости и изменчивости, в котором изменчивость служит его приспособительным реакциям, направленным на поддержание жестких и пластичных констант организма [Анохин П.К, 1975]. Изменение функциональной активности органов и систем, выражающихся в сдвигах пластических констант организма, обеспечивает необходимые условия для выполнения актов жизнедеятельности, а также для поддержания жестких констант.
Первым результатом деятельности управляющих структур является активизация специфических гомеостатических реакций. При невысокой силе раздражителей и изменении внешней и внутренней сред, а также при наличии заранее выработанной адаптации к данному конкретному воздействию адаптационная деятельность ограничивается специфическими гомеостатическими реакциями. Реализация этих, а также любых других актов жизнедеятельности требует энергетического и пластического обеспечения функций. Первоначально эту задачу удается выполнить за счет энергетических возможностей и пластического резерва функционирующих клеток. И эти процессы управления механизмом общей адаптации осуществляются взаимодействием нервных влияний совместно с разными гормональными системами.
В этом плане наиболее подробно изучены особенности функциональной активности эндокринных желез при выполнении различных по длительности и интенсивности физических нагрузок и связь их с тренированностью организма.
Влияние гормонов на обменные процессы при выполнении физических нагрузок, как правило, изучаются при исследовании предельной мобилизации возможностей организма [Haynes R.C. a. all., 1957; Волков Н.И. и соавт., 2000]. Такие исследования имеют большое значение для управления тренировочными нагрузками, процессом восстановления после них и обеспечения развития энергетических, структурных и функциональных основ спортивной работоспособности.
Особое значение в этом аспекте имеют исследования, которые выявили роль моноаминергических систем мозга [Seliy H., 1975; O’Malley B.M., Schroder W.T., 1976]. Показана их роль в двигательной активности животных, изучены и углубляются знания о механизмах их деятельности в процессе поведения, обучения и памяти. Ключом к пониманию этих механизмов является морфо - функциональная концепция о существовании в мозге специфических центров и нейронов в деятельности которых особую роль играют биогенные амины и другие медиаторы.
Гистохимическими и морфологическими исследованиями этих систем раскрываются механизмы, объясняющие связь интенсивности двигательной деятельности с механизмами поведения, эмоциональных реакций.
Показано, что при спортивной тренировке происходят закономерные изменения как в ЦНС, так и эндокринной системе [Кахана М.С., 1970].
ЭЭГ исследования выявили, что очаги возбуждения, прежде всего, возникают в корковых и подкорковых образованиях, а в дальнейшем нервные импульсы распространяются на миндалевидный комплекс, гипоталамус и ростральный отдел ретикулярной формации.
Было показано, что при физических нагрузках нейрогуморальные изменения протекают двухфазно. В результате возбуждения гипоталамических образований в первой фазе усиливается выделение катехоламинов (КА) и гормонов щитовидной железы. Во второй фазе нарастает выделение адренокортикотропного гормона (АКТГ) и кортикостероидов [Баранов Н.Н., Кахана М.С., 1979]. Известно, что структуры мозга, имеющие отношение к функциональной деятельности гипофиза и к мобилизации кортикостероидов, сравнительно богаты КА. Теперь стало известно, что интенсивность нейросекреции коркового и мозгового отделов надпочечников, участвующих в регуляции многочисленных процессов, протекает в тканях мозга не изолированно, а взаимосвязано [Клименко К.С., 1968] и в значительной мере определяется концентрацией адреналина (А), особенно норадреналина (НА) в нервных образованиях гипоталамуса [Комиссаренко В.П., 1968]. В виду того, что основным звеном адаптационных процессов является головной мозг, изучение влияния кортикостероиздых гормонов на ЦНС представляет особый интерес [Митюков М.М., 1968].
Действие физических нагрузок на активность желез внутренней секреции показывает, что в их динамике существенное место занимают ряд факторов - мощность и продолжительность нагрузки, характер физических упражнений и уровень тренированности спортсмена. Перечисленные факторы имеют широкий диапазон воздействия, с одной стороны, а с другой - "отклики" функциональных систем организма имеют еще более широкий индивидуальный диапазон вариативности. И, очевидно, по этой причине выявление общих закономерностей реакций той или иной эндокринной (как и любой другой) системы при тщательном индивидуальном анализе не всегда, но довольно часто входят в противоречие с общегрупповой закономерностью [Jagiełło W. a. all., 2004].
В тех случаях, когда физическая нагрузка не достигает физиологических пределов вариации функциональных характеристик очень велики [Szewczuk B. a. all., 2004; . Tkaczuk W. a. all., 2004].
Описанные состояния не были подвергнуты систематическому анализу и в связи с этим стали предметом данной работы.
2. Материал, методы исследования и их организация
2.1. Материал исследования
В исследования принимали участие 14 квалифицированных борцов классического стиля в возрасте 20,5 ± 0,8 лет. По результатам медицинского освидетельствования все спортсмены по состоянию здоровья были отнесены к здоровым людям.
Стаж тренировки составлял 5 ± 0,5 года.
Для исследования был выбран такой период онтогенеза человека, когда активные росто - весовые изменения почти завершились.
2.2. Методы исследования
Педагогические методы исследования
Исследования проводились в лабораторных условиях.
Функциональное состояние двигательной сенсорной системы определялось с помощью комплексной методики на динохроногониопантографе [Бельхауан М. 1977]. Модельное движение представляло собой воспроизведение (репродукция) прямого угла со сторонами - по абсциссе и ординате по 50 мм. Время выполнения движения составляло 1 секунду при усилии на браншах динамометра в 50 % от максимального возможного для данного спортсмена.
Выбор такого сочетания количественных характеристик движения обусловлен тем, что согласно данным ряда авторов [Мирский М.Л. 1964; Меньшиков В.Я., 1966; Бельхауан М., 1977; Ткачук В., 1986; Czubak K. a. all., 2003] время в 1 секунду, усилие в 50 % от максимального и угол в 90 угловых градусов наиболее точно воспроизводятся спортсменами. Таким путем мы пытались создать наиболее благоприятные условия уменьшающие влияние сбивающих факторов.
Алгоритм работы был следующим:
- серия упражнений в 20 повторений движения,
- перерыв в 3 мин.,
- всего 20 серий (в сумме 400 повторений),
- 17 движений выполнялись под контролем зрительной сенсорной системы,
- 18-20 движения выполнялись без контроля зрения (репродукция на основе проприоцептивной чувствительности).
Перед нагрузкой и после ее окончания забирались порции мочи и крови.
Медико-биологические методы исследования
Одним из наиболее распространенных в эндокринологии методов количественной оценки секреторной активности ЖВС у человека и животных in vivo является определение концентрации гормонов в периферической крови. Этот непрямой способ оценки в виду того, что он отражает не только непосредственную работу ЖВС, но и интенсивность гормонального метаболизма, степень связывая гормонов с белками крови и скорость их секреции. Однако, если функция печени и почек существенно не изменены и уровень плазменных белков, связывающих определенные гормоны в периферической крови может, прямо коррелировать с секреторной активностью соответствующих ЖВС [Комиссаренко В.П., 1968].
Другим адекватным непрямым способом количественной оценки секреторной активности ЖВС является измерение суточной экскреции с мочой гормонов или их специфических метаболитов. Так же в случае нормальной работы печени и почек определение величин экскреции гормональных соединений может пропорционально отражать интенсивность секреторных процессов в соответствующих железах [Комиссаренко В.П., 1968].
В нашей работе использовались оба способа исследования секреторной активности гормонов надпочечников человека.
Для исследования количественных параметров секреции гормонов мозгового слоя надпочечников использовался унифицированный флюорометрический метод определения адреналина (А) и норадреналина (НА) в моче. При анализе применялась методика адсорбции на окиси алюминия с окислением йодом [по Меньшикову В.В., 1961; 1969].
Метод разработан на основе сочетания выделения катехоламинов (КА) на колонке с окисью алюминия [по Euler V.S. van et al., 1959].
Катехоламины определялись в порции мочи по Матлиной Э.Ш. и соавт. [1974], Винницкому В.Б. и соавт. [1978].
Один из гормонов коркового слоя надпочечников - 11-ОКС определяли с помощью унифицированного флюорометического метода определения неконъюгированных 11-ОКС в плазме крови [по Moor P. de et al., 1960], а флюорометический метод определения суммарных неконъюгированных 11-ОКС в моче [по Dorner G., Staki F. 1964].
2.2.3. Методы статистического анализа
Для статистического анализа первичных данных использовался пакет прикладных программ Exsell и Statistica 6.0 [Ferguson G.A., Takane Y. 1997; Шестаков М.П., Попов Г.И., 2002].
2.3. Организация исследований
С целью элиминации циркадной периодики функционального состояния организма спортсменов все исследования проводились в одно и тоже время суток (с 6-00 до 6-30 утра).
Для стабилизации функционального состояния испытуемых были созданы: однообразный режим жизни и питания, количество тренировочных занятий, объем и интенсивность физических нагрузок.
Результаты исследования
Содержание А в моче в среднем по группе обследованных борцов составляло 29,5 ± 5,04 нмоль·сутки-1. При этом индивидуальная вариативность достигала 48,4 %. Величина ошибки при воспроизведении заданного пространства (дифференциального порога проприорецепции) составляла в среднем 4,9 ± 0,4 угловых градуса, что соответствовало абсолютной ошибке в 10,2 % от заданной величины с вариативностью в 25,2 %.
Как видно из представленных количественных показателей вариативность обоих показателей достигает значительных величин. При этом гипотеза о нормальности распределения изучаемой выборки (по критерию χ2) подтвердилась.
Дальнейший анализ был направлен на поиск связи между содержанием А и величиной ошибки при воспроизведении пространства. Линейный корреляционный анализ показал, что между изучаемыми показателями выявлена средняя по тесноте отрицательная по направленности связь (r = - 0, 323 ± 0,11). При этом расчеты свидетельствуют, что вариации величины ошибки при воспроизведении заданного пространства (ОВЗП) только на 10,4% зависят от вариаций А.
Расчет уравнения линейной регрессии позволил получить формулу следующего вида (I):
Yx = 5,1 - 7,1·10-3 x ± 1,1 (I),
где: Yx - прогнозируемая величина ОВЗП, %;
x - содержание А в моче, нмоль·сутки-1
подтвердил установленную связь.
Для проверки гипотезы о линейности связи между изучаемыми признаками был рассчитан коэффициент корреляционного отношения (η) который составил 0,923 ± 0,03. Такой результат свидетельствовал о том, что более адекватно установленная связь может быть описана не линейным, а криволинейным уравнением следующего вида (II):
Yx = - 0,0374 x3+ 0,6667 x2 - 3,5878 x + 9,9543 ± 1,7 (II),
Обозначения те же, что и в уравнении I.
Анализ уравнения II (рис. 1), показал, что между содержанием А и величиной ОВЗП в состоянии относительного физиологического покоя выявлена зависимость, свидетельствующая о том, что точность воспроизведения заданного пространства, определяемая по величине порога различения пространственной характеристики движения, связана с содержанием А в моче. Из представленного уравнения регрессии вытекает, что существует такое содержание А при котором величина порога различения наименьшая, т.е. наблюдается высокая точность воспроизведения заданного пространства. Таким оптимальным содержанием А в моче является зона от 10,9 до 29,5 нмоль·сутки-1.
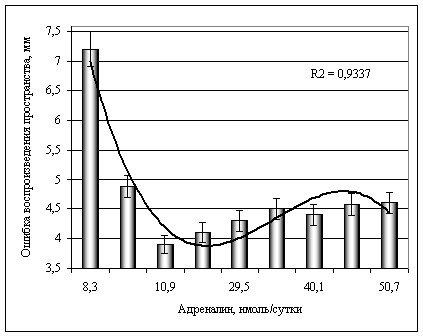
Рис. 1. Зависимость величины ошибки при воспроизведении пространственного параметра движения в состоянии относительного физиологического покоя от концентрации адреналина в моче перед выполнением спортсменами физической нагрузки
Анализ тесноты связи между изучаемыми показателями (по индексу детерминации) показал, что 56,6 % вариаций ОВЗП связаны с изменениями содержания А в моче. Остальные 43,5 % вариаций обусловлены иными экзогенными и эндогенными причинами.
Сразу после окончания операторской работы вновь был проведен анализ содержания А в моче. Оно стало недостоверно меньше на 20,3 %. При этом ОВЗП увеличилась на 18,2 %. Незначительно уменьшилась и теснота связи между ними (r = - 0,319 ± 0,07). Соответственно уменьшился и индекс детерминации (до 10,2 %). Из этого следует то, что только 10,2 % вариаций величины ОВЗП можно связать с вариациями концентрации А в моче.
Преобразилось, в определенной степени, и уравнение регрессии (III):
Yx = 5,39 - 1,71·10-2 x ± 0,52 (III)
(Обозначения те же, что и в уравнении I).
Расчет уравнения адекватно описывающего изучаемые показатели позволил получить полином 3-й степени (IY):
Yx = - 0,0295 x3+ 0,4859 x2 - 2,2868 x + 7,6058 ± 0,01 (IY),
Обозначения те же, что и в уравнении I.
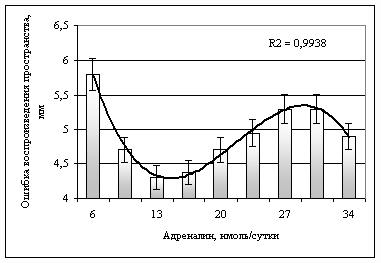
Рис. 2. Зависимость величины ошибки при воспроизведении пространственного параметра движения после физической нагрузки от содержания адреналина в моче перед выполнением спортсменами физической нагрузки
Анализ уравнения IY позволил определить концентрацию А в моче, собранной после нагрузки, при которых величины ошибок при воспроизведении пространственной характеристики движения являются наименьшими. Это диапазон лежит в пределах от 10 до 20 нмоль·сутки-1.
Необходимо обратить внимание на то, что хотя содержание А в моче после нагрузки (монотонная операторская работа) и снизилось по сравнению со состоянием физиологического покоя, оптимум остался таким же как и до работы.
Полученный результат по точности репродукции пространства на 68,1 % зависит от концентрации А в моче после работы.
Следовательно, имеется четкая тенденция, отражающая интенсивность изменений ОВЗП от содержания А в моче и описываемая уравнением регрессии 3-й степени. Суть этих изменений такова, что концентрация А в моче во время монотонной операторской работы нелинейно снижается и в процессе ее снижения имеется диапазон концентрации А в моче при котором точность движения является наилучшей. Этот диапазон лежит в пределах 10 - 20 нмоль·сутки-1. Это значения А не зависят от текущего состояния организма спортсмена (то ли это относительный физиологический покой, то ли операторская работа).
Таким образом, между содержание А в моче и величиной ОВЗП наблюдается сложная функциональная зависимость, свидетельствующая о том, что точность дифференцировки заданного пространства (проприоцептивная чувствительность или порог проприорецепции), а следовательно и функциональное состояния двигательной сенсорной системы, связано с оптимальным содержанием А в моче.
Подобная схема анализа первичного материла исследований была применена и для изучения динамики других гормонов надпочечников - норадреналина (НА), дофамина (ДА) и 11-оксикетостероида (11-ОКС).
Сводные результаты статистической обработки первичных данных приведены в табл. 1.
Таблица 1
Группа уравнений нелинейной регрессии, описывающих взаимодействие различных катехоламинов с величинами ошибок при воспроизведении заданного пространства
|
№ |
Показатели: |
Свобод- |
Коэффи- |
Коэффи- |
Коэффи- |
± m |
|
1. |
НА и ОВЗП (в покое) |
6,0944 |
- 1,5114 |
0,3957 |
-0,0262 |
0,001 |
|
2. |
НА и ОВЗП (после работы) |
3,7008 |
1,7736 |
- 0,5473 |
0,0386 |
0,001 |
|
3. |
ДА и ОВЗП (в покое) |
5,0238 |
0,4442 |
-0,0665 |
- |
0,002 |
|
4. |
ДА и ОВЗП (после работы) |
4,7738 |
3,0446 |
- 0,7745 |
0,05 |
0,001 |
|
5. |
11-ОКС и ОВЗП (в покое) |
11,438 |
- 2,4302 |
0,166 |
- |
0,002 |
|
6. |
11-ОКС и ОВЗП (после работы) |
1,9381 |
1,0339 |
-0,0666 |
- |
0,002 |
Условные обозначения:
НА - норадреналин,
ОВЗП - ошибка воспроизведения заданного пространства,
ДА - дофамин,
11-ОКС - 11-оксикетостероид.
Анализ другого гормона симпатоадреналовой системы (САС) показал, что в состоянии относительного физиологического покоя его содержание в моче (в среднем) составляет 43 ± 5,7 нмоль·сутки-1, а ошибка репродукции пространственного параметра движения составила 5,4º или 11,9 % от заданной величины (90º). Как и А, НА описывается нелинейным уравнением регрессии (табл. 1, 1).
Расчет индекса детерминации свидетельствует, что 62,8 % случаев вариации ОВЗП связаны с изменениями содержания НА в моче и, таким образом, изменения содержания НА коррелируют с уровнем функционального состояния двигательной сенсорной системы.
Под влиянием часовой операторской работы наблюдалось повышение содержания НА в моче на 11,6 % по сравнению с исходным (дорабочим) состоянием. При этом величина ОВЗП снижалась (улучшалась точность репродукции пространственного параметра движения) на 30,4 %.
Высокое значение коэффициента корреляционного отношения (0,845 ± 0,05) давало основание для предположения о нелинейной связи изучаемых показателей. Эта гипотеза подтвердилась при анализе регрессионного уравнения (табл. 1, 2).
Он показывает, что можно выявить две зоны такого содержания НА в моче при которых точность дифференцировки заданного пространства наивысшая (56 - 72 нмоль·сутки-1) и наинизшая (24 - 32 нмоль·сутки-1) для данной группы испытуемых. Такие вариации функционального состояния проприоцептивной сенсорной системы не являются результатом только случайных воздействий на организм. Об этом свидетельствует расчет индекса детерминации, который составил 74,9 %.
Таким образом, функциональное состояние двигательной сенсорной системы тесно связано с вариациями содержания НА в моче спортсменов.
Необходимо подчеркнуть, что в отличие от динамики А, изменения НА после операторской работы имеют противоположную направленность и протекают синхронно, но асинфазно. Варьирует и оптимальное содержание НА в моче в период после работы. Так, если до работы оптимум содержания НА приходился на 25 - 31 нмоль·сутки-1, то после завершения работы он переместился в диапазон 56 - 72 нмоль·сутки-1.
Известно, что при синтезе А и НА используется их предшественник дофамин (ДА). Представляет определенный интерес после анализа динамики А и НА рассмотреть и динамику их предшественника.
Как показал статистический анализ в состоянии относительного физиологического покоя среднегрупповое содержание ДА в моче составило 700 ± 95,2 нмоль·сутки-1. При этом величина ОВЗП равнялась 5,19 ± 0,5º или ошибку в 11,5 % от заданной величины. Коэффициент корреляции, корреляционное отношение и индекс детерминации соответственно были: r = 0,719; η = 1,0; R2 = 0,517.
Эти результаты свидетельствуют, что взаимосвязь изучаемых параметров нелинейная и 51,7 % вариаций ОВЗП коррелируют с содержанием ДА в моче. Представить данную зависимость можно в виде регрессионного уравнения 2-го порядка с приведенными там соответствующими коэффициентами уравнения (табл. 1, 3).
Также как и в динамике А и НА в динамике ДА установлены зона оптимума ( > 1100 нмоль·сутки-1).
Следовательно, функциональное состояние двигательной сенсорной системы связана с динамикой содержания ДА в моче и эти процессы характеризуются нелинейностью. Различие между изучаемыми показателями в состоянии покоя и послерабочим состоянием существует только в степени нелинейности. Так, в состоянии покоя связь может быть адекватно описана уравнением регрессии 2-го порядка, а после работы - 3-го (табл. 1, 4).
Анализ динамики одного из глюкокортикоидных гормонов - 11-ОКС показал, что в среднем его содержание в моче составило 700 ± 91,3 нмоль·сутки-1 при ОВЗП в 5,12º или 11,4 % от заданной величины.
Корреляционный анализ выявил тесную отрицательную связь (- 0,736 ± 0,09).
Коэффициенты регрессионного уравнения как в состоянии покоя, так и после работы приведены в табл. 1, 5-6.
Полученные данные позволяют отметить, что часовая монотонная операторская работа изменяет характер связи между содержанием 11-ОКС и ОВЗП на противоположный, а по мере увеличения содержания гормона точность воспроизведения пространственного параметра движения снижается.
Дискуссия
Рассмотрев раздельно динамику ОВЗП в зависимости от различных гормонов САС представляет интерес провести сопоставление их совместной динамики. Такой анализ позволит выявить как функциональное состояние проприоцептивной сенсорной системы человека связано с содержанием гормонов коры надпочечников в моче [Ткачук В.Г., 1986]. Как вытекает из экспериментальных данных, представленных на рис. 3, из всего многообразия вариаций ОВЗП можно выделить следующие типичные варианты:
- синхронные синфазные колебания функционального состояния двигательной сенсорной системы (А и НА в состоянии покоя),
- синхронные асинфазные колебания (НА до и после работы, 11-ОКС в моче до и после работы),
- асинхронные колебания - (ДА до и после работы, А - до и после работы) [Ткачук В.Г., 1986; W. Jagiello a. all., 2003].
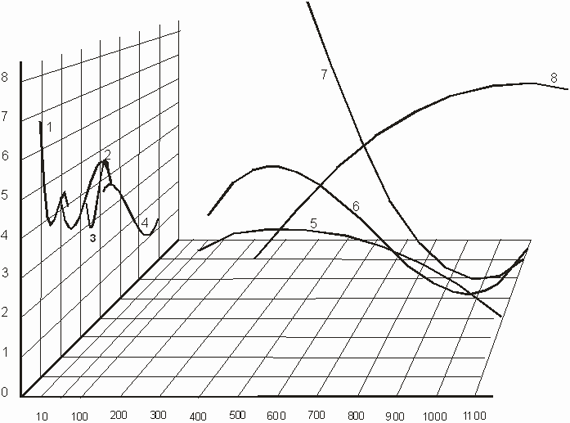
Рис. 3. Графики уравнений регрессии различных гормонов
Условные обозначения:
ОВЗП - ошибка воспроизведения заданного пространства, угл. градусы;
1 - адреналин до работы,
2 - адреналин после работы;
3 - норадреналин до работы,
4 - норадреналин после работы,
5 - дофамин до работы,
6 - дофамин после работы,
7 - 11-ОКС до работы,
8 - 11-ОКС после работы).
Все без исключения установленные связи имеют выраженные тенденции, которые можно описать с помощью регрессионных моделей . Эти модели дают возможность судить о динамике изучаемого процесса и прогнозировать направленность развития функционального состояния двигательной сенсорной системы на достаточно длительном интервале времени. Локальные колебания ОВЗП более точно описываются регрессионными моделями 2-го и 3-го порядков.
Их анализ позволяет в зависимости от содержания в моче того или иного гормона надпочечников определить фазы повышенной и пониженной сенсорной чувствительности. И эти фазы имеют существенное значение как при выполнении точностных движений во время тренировок и соревнований, а особенно при обучении новым точностных двигательным действиям. Их важность вытекает из того факта, что в фазу повышенной чувствительности сенсорной системы ошибка при выполнении пространственной компоненты движения уменьшается с 20 - 25 % от заданной величины до 5 - 7 %.
Известно, что изучение реакций целостного организма человека, с одной стороны, дает определенные преимущества в понимании действующих при этом механизмов управления функциями, но, с другой стороны, вводит целую систему определенных ограничений.
Так "...ни механическое соединение кости, крови, мускулов, тканей и т.д., ни химическое соединение элементов не составляют еще живого" "...организм есть, несомненно высшее единство, связывающее в себе в одно целое механику, физику и химию, так что эту троицу нельзя больше разделить" [Маркс К., Энгельс Ф., 1954]. И как справедливо отмечает Афанасьев В.Г. [1980] "...понятие целого, целостной системы отражает одну из важнейших черт предметов и явлений материальной действительности: их способность вступать в такого рода в з а и м о д е й с т в и я (разрядка наша), в результате которых образуются новые качества, не присущие исходным объектам взаимодействия. Отсюда вытекает, что категория целостности весьма близка к категории качества, ибо качество есть известное выражение целостного предмета - одна из его качественных характеристик" (с. 29).
Исходя из такой теоретической предпосылки в данной работе и были объединены все три перечисленные особенности - изучение целостного организма, взаимодействие его подсистем между собой и организма со средой, и качественные показатели деятельности двигательной сенсорной системы.
Проведенные и представленные выше результаты исследований позволили сделать следующие выводы.
Выводы
1. Вариативность функционального состояния двигательной сенсорной системы является закономерным следствием суммации внутри- и межсистемных взаимосвязей случайных и детерминированных колебательных процессов, происходящих в центральной нервной, двигательной и эндокринной системах организма человека.
2. Динамика функционального состояния двигательной сенсорной системы при воспроизведении основных характеристик движения аппроксимируется нелинейными регрессионными моделями, свидетельствующими о том, что точность дифференцировки характеристик движения (пространство, время, усилие) волнообразно изменяется при изменении содержания гомонов в моче и крови.
3. Наиболее информативным показателем функционального состояния двигательной сенсорной системы при выполнении движений, требующих высокой точности является уровень дофамина в урине.
4. Физиологическая сущность вариативности заключается в ее роли как одного из многочисленных механизмов осуществления долговременной адаптации систем управления произвольными и непроизвольными движениями человека в процессе обучения, а также при срочной адаптации в процессе конкретной деятельности и текущего состояния организма. Она (вариативность) является одним из элементов творческих (бессознательных) функций приспособления к динамичным условиям внешней и внутренней среды.
Литература
3. Анохин П.К., Биология и нейрофизиология условного рефлекса. - М.: Медицина, 1868. - 548 с.
4. Анохин П.К. Очерки по физиологии функциональных систем. - М.: Медицина, 1975. - 448 с.
5. Афанасьев В.Г. Системность и общество. - М.: Политиздат, 1980. - 368 с.
6. Баранов Н.Н., Кохана М.С. Нейрогормональные механизмы тренированности. - Кишенев: Штиинца, 1979. - 120 с.
7. Бельхауан М. Моделирование соотношения параметиров двигательной координации у гимнастов // Моделирование функционального состояния спортсменов различной подготовленности (Сборник научных работ). - Киев, 1977, с. 51-56.
8. Винницкий В.Б., Шмалько Ю.П. К методике определения адреналина, норадреналина, ДОФА и дофамина в одной порции мочи // Лабораторное дело, 1978, № 1. - С. 688 - 690.
9. Виру А.А., Кырге П.К. Гормоны и спортивная работоспособность. - М.: Физкультура и спорт, 1983. - 159 с.
10. Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности. Киев, Олимпийская литература. 2000. - 503 с.
11. Кахана М.С. Роль гипоталамуса в механизме гомеостаза желез внутренней секреции // Гипоталамо-эндокринные взаимоотношения. - Кишенев, 1970, вып. 2, С. 3 - 47.
12. Клименко К.С. Роль гормонов коркового и мозгового слове надпочечников в регуляции дыхательной активности ткани головного мозга // Гормоны и головной мозг: Наукова думка, Киев, 1968. - С. 103 - 106.
13. Комиссаренко В.П. Гормоны и мозг // Гормоны и головной мозг: Наукова думка, Киев, 1968. - С. 5 - 13.
14. Маркс К., Энгельс Ф., 1954, Собр. соч., т. 20.
15. Матлина Э.Ш., Киселева З.М., Софиева И.З. Метод определения адреналина, норадреналина, дофамина и ДОФА в одной порции мочи // Методы клинической биохимии гормонов и медиаторов. Ч. 2. - М.: 1-й МОЛМИ, 1974. - С. 27.
16. Меерсон Ф.З. Пластическое обеспечение функций организма. - М.: Наука, 1967. - 318 с.
17. Меньшиков В.В. Об определении катехоламинов в моче // Лабораторное дело, 1961, № 4. - С. 18 - 21.
18. Меньшиков В.В. Методы клинической биохимии гормонов и медиаторов. - Учебное пособие, 2-е изд. - М: 1-й Моск. мед. инст., 1969. - 160 с.
19. Меньшиков В.Я. Исследование способности дифференцировать мышечное напряжение, пространство и время у юных гимнастов : Автореф. Дисс...канд. пед. наук. - М., 1966. - 24 с.
20. Мирский М.Л. Исследование координации мышечных напряжений при дозировании усилий : Автореф. дисс...канд. биол. наук. - М., 1964. - 20 с.
21. Митюков М.М. Гормоны коры надпочечников и реакции нервной системы на стрессорные воздействия // Гормоны и головной мозг: Наукова думка, Киев, 1968. - С. 24 - 30.
22. Митюков М.И., Богданов Т.С., Горина И.Л., Емельянов Н.А., Ноздрачев А.Д., Подвигина Т.Т., Ракицкая В.В., Соколова Е.В., Шаляпина В.Г. Гормоны коры надпочечников и центральная нервная система. - Л.: Наука, 1970. - 160 с.;
23. Невядомский М.М. Медицинская мысль Узбекистана, 1927, № 5, с. 15 - 17
24. Розен В.Б. Основы эндокринологии. - М.: Высшая школа, 1980. - 344 с.
25. Ткачук В.Г. Механизмы вариативности при управлении точностными движениями человека. Дисс...док. биол. наук, специальность- управление в биологических и медицинских системах. Киев. Ин - т Кибернетики им. В. Глушкова НАН Украины, 1986. - 382 с.
26. Шестаков М.П., Попов Г.И. Статистика (Обработка спортивных данных на компьютере) // М.: СпортАкадемПресс. - 2002. - 278 с.
27. Яковлев Н.Н. Биохимия спорта. - М.: Физкультура и спорт, 1974. - 288 с.
28. Czubak K., Ignasiak D., Tkaczuk W. Computeruntersuchung der bewegungskoordination der sportler. Gemeinsames Kolloquium der Bewegungs - und Trainingswissenschaft des Institutes fur Sportwissenschaft, vom. 13. bis 14. Oktober 2003, str. 51.
29. Dorner G., Staki F. Fine einfache fluorometrische methode zur routinebestimmung von fralem cortisol + corticosteron im harm // Acta biol. Et med/ ger., 1964. Bd. 12H.5. - S. 600 - 611.
30. Euler V.S. van, Lishejko F. Estimation of catecholamines in urine // Acta Physiol. Scand., 1959, v. 27. - P. 122 - 132.
31. Ferguson G.A., Takane Y. Analiza statystyczna w psychologii i pedagogice. PWN. Warszawa, 1997.
32. Jagiello W., Tkaczuk W., Jagiello M. Zmiany możliwości siłowych w rocznym szkoleniu kwalifikowanych tenisistów. XII Międzynarodowa Konferencja Naukowo-Metodyczna "Kierunki doskonalenia treningu i walki sportowej", COS Spała, 1-3 XII 2003, str. 91-92.
33. Jagiello W., Kalina R.M., Tkaczuk W. Development of strength abilities in children and youths. Biology of Sport, Vol. 21, Nr 4, 2004, S. 351-368.
34. Greep, R.O., Astwood, E.B. (editors): Handbook of physiology. Section 7, endocrinology, the adrenal gland, Baltimore, 1975. The Williams and Wilkins Co., vol. 6.
35. Haynes R.C., Jr., Berthet L. Studiees on the mechanism of adrenocorticotropic hormone. J. Boil. Chem. 225: 115, 1957.
36. Moor P. de, Steeno O., Raskin M., Hendrix A. Fluorimetric determination of the plasma 11-hydroxycorticosteroids im man // Acta endokrinol., 1960, v. 33. - P. 297 - 307.
37. O'Malley B.W., Schrader W.T. The receptors of steroid hormones, Sci. Am. 234:32, 1976.
38. Selye H. Confusion and controversy in the stress field, J. Hum. Stress, 1:37, 1975.
39. Greep R.O., Astwood E.B. (editors): Handbook of physiology. Section 7, endocrinology, the adrenal gland, Baltimore, 1975. The Williams and Wilkins Co., vol. 6.
40. Haynes R.C., Jr., Berthet L. Studiees on the mechanism of adrenocorticotropic hormone. J. Boil. Chem. 225: 115, 1957.
41. Moor P. de, Steeno O., Raskin M., Hendrix A. Fluorimetric determination of the plasma 11-hydroxycorticosteroids im man // Acta endokrinol., 1960, v. 33. - P. 297 - 307.
42. O'Malley B.W., Schrader W.T. The receptors of steroid hormones, Sci. Am. 234:32, 1976.
43. Selye H. Confusion and controversy in the stress field, J. Hum. Stress, 1:37, 1975.
44. Szewczuk B., Tkaczuk W., Jagiello W., Petrowicz B., Skowron J., Ojrzanowski A. Solving the problems of athletes’ functional state control by means of data net processing. Sport training in interdisciplinary scientific researches. Częstochowa, 2004. - S. 78-83.
45. Tkaczuk W., Rowny A., Jagiello W. Assessment of sensory functional system In managing movement. UGDYMAS. KUNO. KULTURA. SPORTAS. Education Physical Training Sport. Lithuania Academy of Physical Education. № 1, 2004. - С.57-67.
Поступила в редакцию 20.05.2005г.